

You are here: Home > Section on Molecular Neurobiology
Neuregulin-ErbB Signaling in Neuronal Development and Psychiatric Disorders

- Andres Buonanno, PhD, Head, Section on Molecular Neurobiology
- Detlef Vullhorst, PhD, Staff Scientist
- Irina Karavanova, PhD, Biologist
- Robert Mitchell, PhD, Intramural Research Training Award Fellow
- Swagata Roychowdhury, PhD, Intramural Research Training Award Fellow
- Miguel Skirzewski, MS, Visiting Fellow
- Carolyn Keating, BS, Postbaccalaureate Fellow
- Katrina E. Furth, BS, Pre–Intramural Research Training Award Fellow
Failure of cortical microcircuits to properly regulate excitatory/inhibitory (E/I) balance is a key feature in the etiology of several developmental psychiatric disorders and neurological diseases, such as schizophrenia, autism, ADHD, and epilepsy. E/I balance is important to synchronize the firing pattern of local neuron ensembles, and its dysregulation can degrade cognitive functions and, in extreme cases, result in epileptiform activity. Network activity, in particular oscillatory activity in the gamma-frequency range (30–80 Hz), is altered in psychiatric disorders and may account for their cognitive and behavioral symptoms. We are interested in how Neuregulin and its receptor ErbB4, which are both genetically linked to psychiatric disorders and function in rodents in an activity-dependent fashion (i.e., experience) in the developing brain to regulate synaptic plasticity, neuronal network activity, and behaviors that model features of psychiatric disorders. We identified a functional interaction between Neuregulin/ErbB4, GABAergic, and dopamine signaling in GABAergic interneurons that is critical for understanding how Neuregulin (NRG) can regulate E/I balance and synchronous activity in neuronal networks, processes that are important for cognitive functions altered in psychiatric disorders.
Our earlier studies demonstrated that, in the hippocampus and neocortex, expression of ErbB4, the major NRG neuronal receptor, is restricted to GABAergic interneurons, particularly the parvalbumin-positive (PV+) fast-spiking interneurons that are critical for modulating gamma oscillation induction and strength (i.e., power). Using genetically targeted mouse mutant models, we went on to show that NRG-ErbB4 signaling regulates synaptic plasticity, neuronal network activity, and behaviors associated with psychiatric disorders. More recently, our group has been investigating other aspects of Neuregulin expression throughout the brain, its processing in response to neuronal activity, and its function in distinct neuronal populations of the developing and maturing nervous system. To achieve these goals, we are using a combination of techniques, including electrophysiological recordings in acute brain slices prepared from normal and genetically altered mice, multi-electrode field recordings from brains of freely moving rats, reverse-microdialysis neurochemistry, confocal fluorescence microscopy in fixed and live tissue, proteomics analyses, and behavioral testing. The ultimate goal of this multi-disciplinary approach is to generate holistic models to investigate the developmental impact of genes that modulate E/I balance and neuronal network activity, and which consequently affect behaviors and cognitive functions altered in psychiatric disorders.
Functional analysis of the extra-synaptic ErbB4 receptor proteome using proteomics approach; identifying an interaction with GABA-A receptors
Most studies on NRG-ErbB4 signaling have focused on the receptor that accumulates at glutamatergic post-synaptic densities (PSDs). However, little research has been performed on the pool of extra-synaptic ErbB4 receptors that may constitute up to 80% of the total receptor population in cortical and hippocampal interneurons. In collaboration with Sanford Markey's group, our laboratory used ErbB4 immunoprecipitation from the soluble fraction of metabolically active synaptosomes followed by LC/MS/MS (liquid chromatography/mass spectrometry/tandem mass spectrometry) to characterize the ErbB4 proteome. Using this approach, we identified the GABA-A receptor α1 subunit (GABAR α1) as an ErbB4–interacting protein. Consistent with this observation, we found that GABAR α1 receptors are highly expressed in ErbB4–positive fast-spiking PV+ in the hippocampus (Figure 1) and that ErbB4 knockout mice have reduced GABAR α1 levels. Remarkably, we found that this novel ErbB4 signaling pathway, which suppresses postsynaptic GABAR currents on GABAergic interneurons, acts independently of ErbB4s canonical receptor tyrosine kinase (RTK) activity. While the effects of NRG on GABAR α1 internalization do not require RTK activity (Figure 1), the ErbB4 protein is necessary for clathrin-mediated endocytosis and reduction of GABA receptor IPSCs (inhibitory postsynaptic currents) (1).
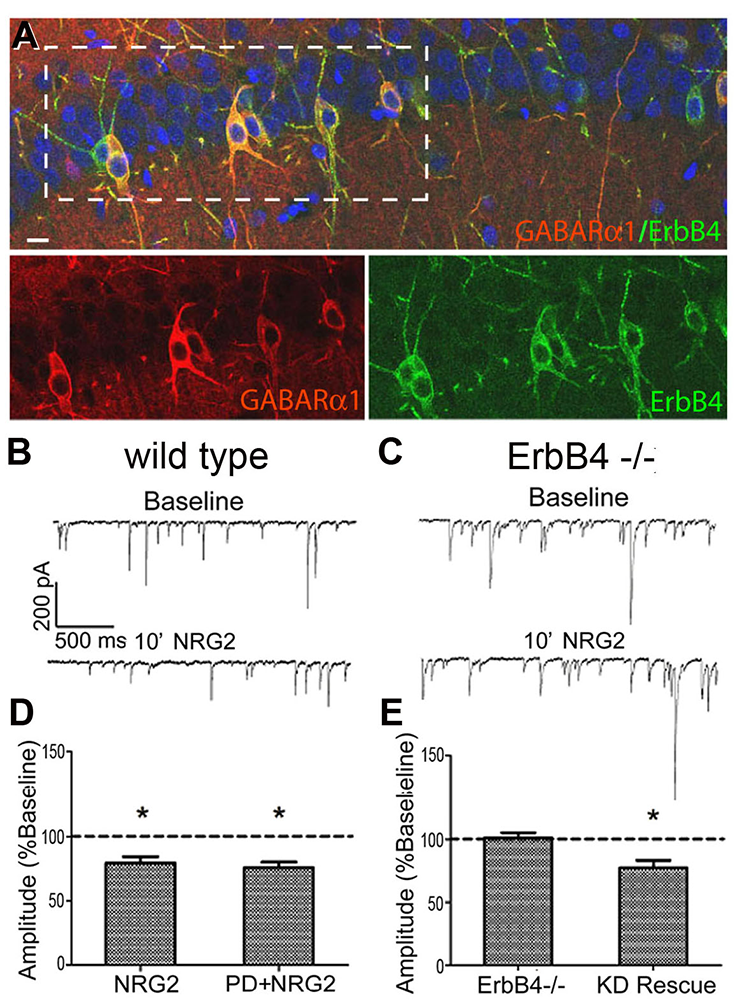
Click image to enlarge.
Figure 1. GABARα1 interacts with ErbB4 to reduce postsynaptic GABAR currents on GABAergic interneurons.
A: GABARα1 and ErbB4 are co-expressed in CA1 hippocampal interneurons. B: Whole-cell voltage-clamp mIPSC recording from wild-type (wt) ErbB4+ interneuron at baseline and following NRG2 application. C: Same as B but the recording is from an ErbB4−/− interneuron. D: mIPSC amplitude analysis of wt hippocampal interneurons treated with NRG2 and NRG2 + the ErbB kinase inhibitor PD158780, indicating that the NRG2 effect does not require receptor tyrosine kinase (RTK) activity. E: Summary of mIPSC amplitude following NRG2 treatment of either ErbB4−/− cultures or ErbB4−/− cultures infected with kinase-dead (KD) ErbB4 AAV vector.
Importance of Neuregulin/ErbB4 and dopamine (DA) signaling in parvalbumin-positive fast-spiking interneurons
Using reverse-microdialysis neurochemistry and electrophysiology, we reported that NRG1 potently triggers dopamine (DA) release in the hippocampus, reverses long-term potentiation (LTP) at Schaeffer collateral-to-CA1 (SC-CA1) synapses and regulates the power of kainate-induced (KI) gamma oscillations in acute hippocampal slices through activation of DA D4 receptors (D4Rs). Importantly, we found that the ErbB4 receptor is not detectable in excitatory pyramidal neurons but is abundantly expressed in GABAergic interneurons, in particular PV+ fast-spiking basket cells. These observations strongly suggested that effects of NRG/ErbB4 on SC-CA1 synaptic plasticity and KI gamma oscillations results from an interplay between the glutamatergic and DAergic neurotransmitter systems. First, we addressed the role of ErbB4 receptors specifically in PV+ GABAergic interneurons by comparing the effects of full vs. interneuron-specific Erbb4 ablation in genetically engineered mice (Figure 2). Unexpectedly, ErbB4 ablation in PV+ interneurons was sufficient to reproduce the effects of the full ErbB4 knockout on LTP. Because our earlier studies found that D4Rs are necessary to mediate the effects of ErbB4 on LTP, experiments are in progress to determine the role of D4Rs in PV+ interneurons. This is a particularly interesting question because, as our laboratory previously reported, a subset of PV+ fast-spiking basket cells co-express D4 and ErbB4 receptors that modulate KI gamma oscillation power in acute hippocampal slices (Andersson RH et al., Proc Natl Acad Sci USA 2012;109:13118). The novel cross-talk between D4 and ErbB4 receptor signaling to augment gamma oscillation power suggests that PV+ interneurons are a critical nexus between NRG/ErbB4 and DA co-signaling and provides further evidence in support of a possible contribution of this pathway to the pathophysiology underlying psychiatric disorders (2,3).
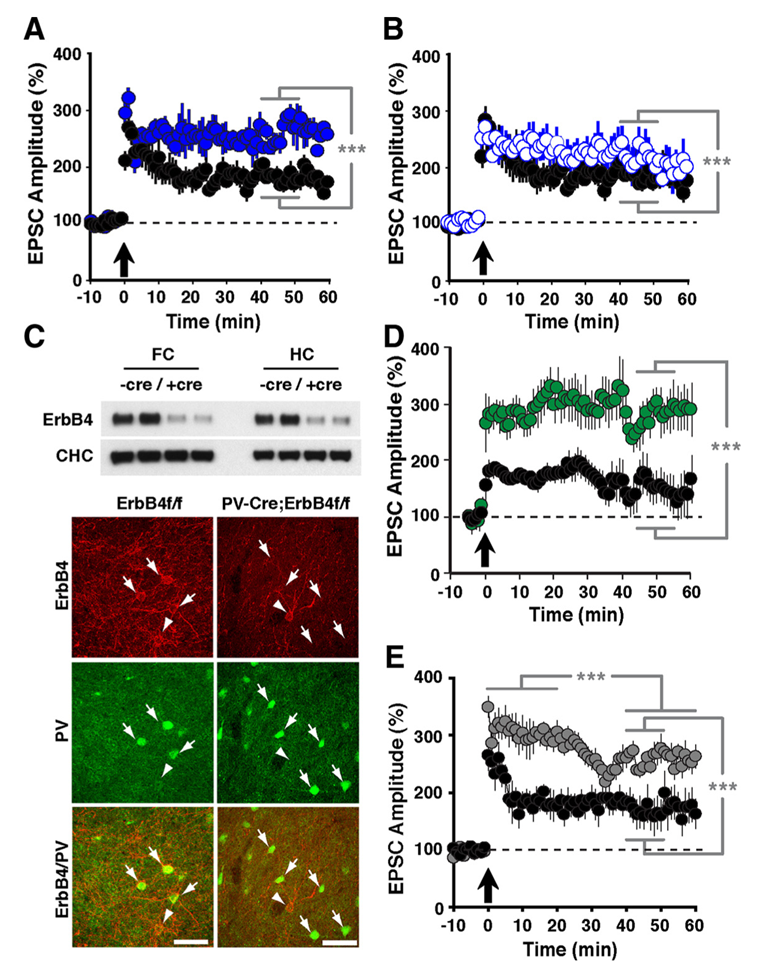
Click image to enlarge.
Figure 2. LTP in ErbB4– and NRG-1–mutant mice
Compared with normal controls (black circles in A, B, D, and E), slices from ErbB4−/− (A: solid blue circles) and ErbB4−/+ mice (B: open blue circles) exhibited enhanced LTP following pairing. C: Top, Western blot analysis of ErbB4 protein in whole-tissue lysates from frontal cortex (FC) and hippocampus (HC) of control ErbB4f/f (−Cre) and PV-Cre;ErbB4 (+Cre) mice. Representative results from two animals are shown for each genotype. Bottom, co-immunofluorescence of ErbB4 (red) and PV (green) in the frontal cortex. In ErbB4f/f controls, numerous PV+ cells coexpress ErbB4 (arrows). In contrast, ErbB4 is not detected in PV+ cells from PV–Cre;ErbB4 mice. Arrowheads mark ErbB4+ cells that are negative for PV. D: Selective ablation of ErbB4 in PV interneurons (green) results in higher LTP. E: LTP was also elevated in slices from Nreg-1+/− mice (grey).
Mesocortical and nigrostriatal DA function in mice with targeted mutations of ErbB4 in PV+ and tyrosine hydroxylase-positive (TH+) neurons
Dysfunctional Neuregulin–ErbB4 signaling in the hippocampus, pre-frontal cortex (PFC), and striatum may contribute to alterations in DA function associated with several schizophrenia symptoms. Because we have shown that NRG1 acutely increases extracellular DA levels to regulate LTP and gamma oscillations, and that ErbB4 expression is confined to GABAergic interneurons (cortex) and TH+ to mesocortical DA neurons, we are investigating the relative role NRG/ErbB4 signaling in the two different neuronal populations. In collaboration with Alon Shamir, we are thus measuring basal extracellular DA levels in the PFC, hippocampus, and striatum in mice harboring targeted mutations of the receptor in either PV+ or TH+ neurons (Skirzewski et al., in preparation). We also are comparing their behaviors to begin unraveling the role of Neregulin-ErbB4 signaling in PV+ interneurons vs. TH+ terminals in regulating schizophrenia-associated behaviors (4).
Dopamine regulation of prefrontal cortical gamma oscillations in freely moving rats
Mounting evidence suggests that gamma oscillations are atypically high at baseline in disorders that affect attention such as schizophrenia and ADHD. A reduced evoked gamma oscillatory power in these subjects, compared with healthy controls, may constitute an endophenotype associated with risk for developing the psychiatric disorders. Based on these observations, we hypothesized that drugs targeting either the D4 or ErbB4 receptors may have therapeutic potential in treating cognitive deficits in patients with schizophrenia. To examine abnormal oscillatory patterns and connect them with the firing patterns of single neurons, we in collaboration with Judith Walters are using multi-electrode recordings from the medial prefrontal cortex and dorsomedial thalamus of rats acutely treated with ketamine to analyze the effects of D4– and ErbB4–targeting drugs on gamma oscillations in this mouse model with "face validity" for schizophrenia (Figure 3).
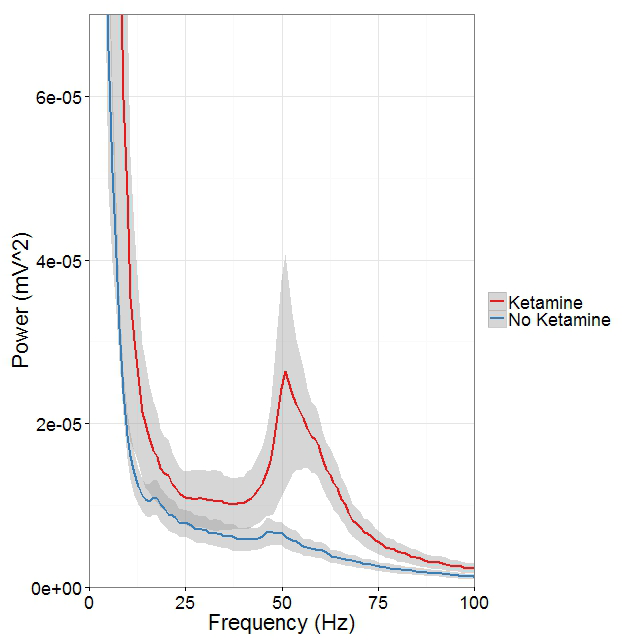
Click image to enlarge.
Figure 3. Ketamine raises gamma (40-65Hz) frequency power in the mPFC during controlled movement.
Multi-electrode recordings in the mPFC revealed an over three-fold increase in gamma (40–65Hz) frequency power while rats walked counter-clockwise at 9–10 revolutions per minute on a circular treadmill. Blue traces come from an epoch 30 minutes before ketamine administration, and the red trace comes from epochs 15 minutes after ketamine administration.
A new paradigm for Neuregulin signaling in the CNS
Little is known about the cellular and molecular processes that promote the conversion of NRG ligands from inactive pro-forms into signaling-competent ligands that can engage ErbB4 receptors to mediate their aforementioned biological effects in the developing and maturing brain. To address this major unresolved question, we investigated the role of NRG2, a NRG isotype that is prominently expressed in the developing postnatal and adult CNS. Traditionally, cortical ErbB4 signaling has been viewed as mediated primarily by axonally derived NRGs from glutamatergic and cholinergic inputs. Consistent with this notion, NRG2 mRNA has previously been reported in dentate gyrus granule cells. However, using a novel technique for double-labeling in situ hybridization called RNAScope (Figure 4), as well as new monoclonal antibodies developed in our lab, we found that, in the rodent hippocampus, NRG2 mRNA and protein are also expressed in ErbB4–positive GABAergic interneurons, suggesting that NRG2 engages in both autocrine and paracrine ErbB4 signaling (Figure 5). Interestingly, we found no evidence of NRG2 protein in axons or axon terminals. Instead, NRG2 accumulates on the soma and proximal dendrites, where it forms large puncta on the plasma membrane that typically do not colocalize with pre- or post-synaptic markers. Rather, immunogold electron microscopy of NRG2 in dissociated hippocampal neurons, performed in collaboration with Susan Cheng, revealed that these puncta are frequently found atop subsurface cisterns, sites of close apposition between the plasma membrane and the endoplasmic reticulum (Figure 5). Double-immunofluorescence labeling of NRG2 with antibodies against the extra- and intracellular domains indicates that the clusters consist of the unprocessed pro-form (Vullhorst et al., submitted). Taken together, the data suggest that NRG2 subserves distinct and non-overlapping roles. We hypothesize that clustering of pro-NRG2 serves to protect the protein from constitutive processing by extracellular sheddases and to render the process of ectodomain shedding subject to some form of regulation by neural signals.
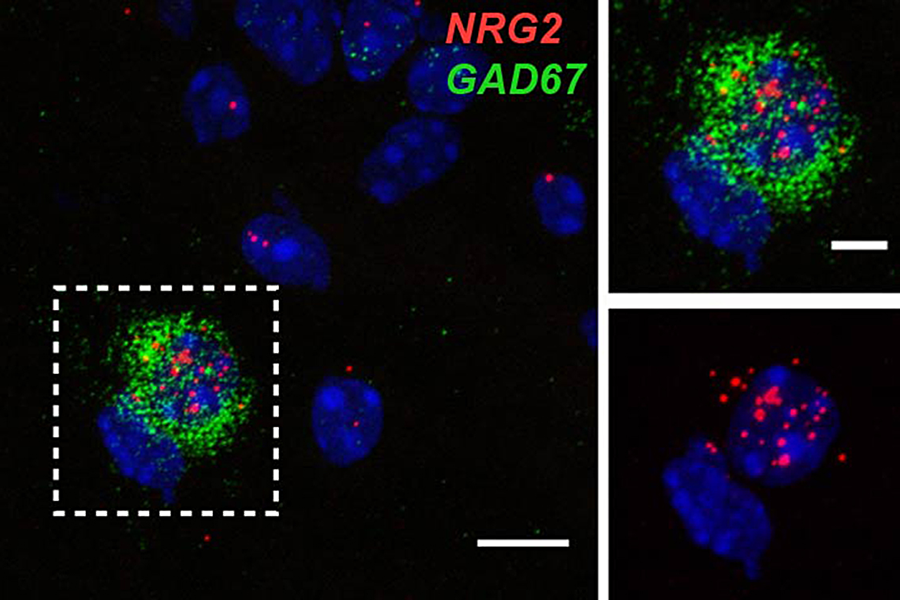
Click image to enlarge.
Figure 4. RNAScope double-label in situ hybridization shows NRG2 and GAD67 co-expression in hippocampal interneurons.
Double immunofluorescence in situ hybridization of NRG2 (red) and Gad67 (green) shows overlapping signals in a neuron located in the stratum oriens of area CA1. DAPI was added to label nuclei (blue). Boundary box on the left outlines the area magnified on the right.
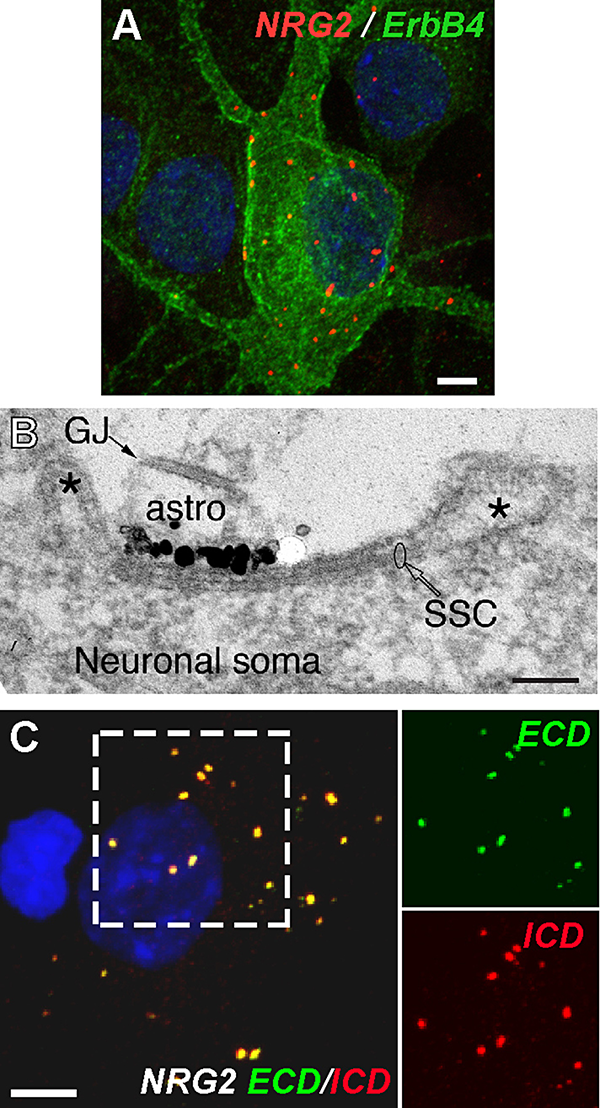
Click image to enlarge.
Figure 5. Neuregulin 2 accumulates at subsurface cisterns of ErbB4–expressing GABAergic interneurons.
A: Cultured hippocampal interneurons co-express NRG2 and its receptor ErbB4. B: Immunogold EM of NRG2 shows a dense signal cluster on the plasma membrane atop a subsurface cistern (SSC). Astrocytic processes (astro, identified by a gap junction [GJ] between two processes) are often closely associated with SSCs. C: Extensive overlap of extracellular (ECD) and intracellular (ICD) domain signals indicate that somatodendritic NRG2 clusters are unprocessed.
Additional Funding
- NICHD DIR Director's Investigator Awards (RRC # D-14-07)
Publications
- Mitchell RM, Janssen MJ, Karavanova I, Vullhorst D, Furth K, Makuskey A, Markey S, Buonanno A. ErbB4 receptor reduces synaptic GABAA currents independently of its tyrosine kinase activity. Proc Natl Acad Sci USA 2013;110:19603-19608.
- Furth KE, Mastwal S, Wang KH, Buonanno A, Vullhorst D. Dopamine, cognitive function, and gamma oscillations: role of D4 receptors. Front Cell Neurosci 2013;7:102 (1-19).
- Penzes P, Buonanno A, Passafaro M, Sala C, Sweet RA. Developmental vulnerability of synapses and circuits associated with neuropsychiatric disorders. J Neurochem 2013;126:165-182.
- Golania I, Tadmorb H, Buonanno A, Kremer I, Shamir A. Disruption of the ErbB signaling in adolescence increases striatal dopamine levels and affects learning and hedonic-like behavior in the adult mouse. Eur Neuropsychopharmacol 2014;in press.
- Vizi ES, Buonanno A. Extrasynaptic ionotropic receptors. Brain Res Bull 2013;93:1-3.
Collaborators
- Jung Hwa (Susan) Cheng, PhD, EM Facility, NINDS, Bethesda, MD
- Catherine Fenster, PhD, The College of Wooster, Wooster, OH
- Luis Hernández, MD, Universidad de los Andes, Mérida, Venezuela
- Ilana Kremer, MD, The Ruth and Bruce Rappaport Faculty of Medicine, Technion–Israel Institute of Technology, Haifa, Israel
- John Lisman, PhD, Brandeis University, Waltham, MA
- Sanford P. Markey, PhD, Laboratory of Neurotoxicology, NIMH, Bethesda, MD
- Jörg Neddens, PhD, JSW Life Sciences, Grambach, Austria
- Alon Shamir, PhD, Mazra Mental Health Center, Akko, Israel
- Judith R. Walters, PhD, Experimental Therapeutics Branch, NINDS, Bethesda, MD
Contact
For more information, email buonanno@mail.nih.gov or visit smn.nichd.nih.gov or neuroscience.nih.gov/Faculty/Profile/andres-buonanno.aspx.