



You are here: Home > Section on Neuroendocrinology
Pineal Gland, Chronobiology, and Neuroepigenetics

- David C. Klein, PhD, Head, Section on Neuroendocrinology
- Diego Bustos, PhD, Postdoctoral Fellow
- Sam Clokie, PhD, Postdoctoral Fellow
- Steven L. Coon, PhD, Staff Scientist
- Surajit Ganguly, PhD, Guest Researcher
- M.A.A. Namboodiri, PhD, Guest Researcher
- Margaret Ochocinska, PhD, Postdoctoral Fellow
- Jiri Pavlicek, PhD, Postdoctoral Fellow
- Harvey Pollard, MD, PhD, Guest Researcher
- Martin Rath, Special Volunteer
- Louise Rovsing, Special Volunteer
- Joan L. Weller, BA, Senior Research Assistant
The Section on Neuroendocrinology is dedicated to the study of the pineal gland and regulation of the pineal hormone melatonin. The work has broad implications for vertebrate biology and is of special interest to clinical scientists studying human diseases relating to circadian rhythms, including endocrine pathologies, sleep and mood disorders, and deficiencies in alertness. The Section also addresses the factors that control the remarkable global changes in gene expression occurring in the pineal gland on a 24-hour basis as well as factors associated with the establishment of the pineal phenotype.
The timezyme: arylalkylamine N-acetyltransferase
A pivotal advance in our investigations into arylalkylamine N-acetyltransferase (AANAT), the second enzyme in the pathway for melatonin synthesis from serotonin, was our finding that the enzyme is critical to the control of the rhythm in melatonin synthesis. In all species examined to date, the large increase in melatonin synthesis at night causes an increase in the production of melatonin, which is a highly conserved feature of vertebrate physiology.
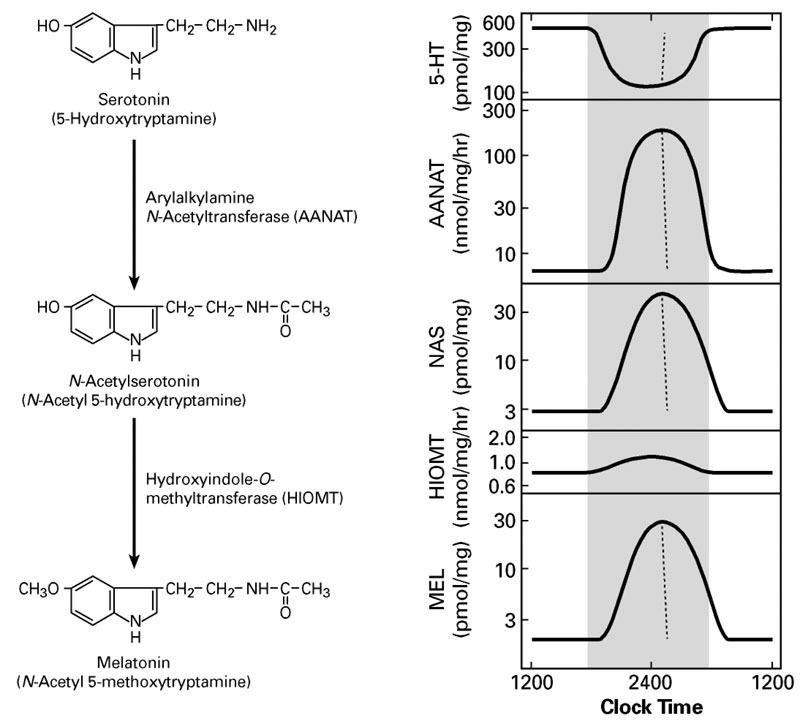
Click for a larger version.
Figure 1.
Regulation of pineal function
The transcriptional mechanisms that control expression of the AANAT gene include interactions of cyclic AMP–response elements in the promoter region; other response elements in the promoter, including the E-box and photoreceptor conserved elements, appear to determine the marked tissue-specific pattern of expression of the gene, which is limited to the pineal gland and retina. We determined that, in rodents, expression of AANAT rises about 100-fold at night; however, this feature of regulation is not highly conserved. We discovered little or no nocturnal increase in AANAT expression in ungulates and primates. Thus, transcriptional control mechanisms are not essential for regulating melatonin synthesis in all vertebrates.
With regard to post-translational control, sequence analysis revealed that all vertebrate AANATs contain C- and N-terminal sites for phosphorylation by cyclic AMP–dependent protein kinase. We determined that the sites control the stability and biological half-life of the enzyme. Phosphorylation of the sites controls binding of AANAT to 14-3-3 protein, which protects the enzyme from destruction; when the enzyme is released, it can be destroyed rapidly. The phosphorylation state of AANAT and binding to 14-3-3 appear to govern the balance between destruction and protection. In collaboration with Fred Dyda and Alison Hickman, we determined the molecular basis of this interaction by analyzing the crystal structure formed by AANAT and 14-3-3 proteins. In collaboration with Benoit Malpaux and Philippe Chemineau, we demonstrated that the interaction is critical for regulation of the enzyme in vivo (Ganguly et al., Proc Natl Acad Sci USA 2005;102:1222). In collaboration with Alastair Aitken, we used a comparative approach, based on both mRNA and protein analysis, to analyze and establish the developmental appearance of each 14-3-3 isoform in the pineal gland and found marked differences in the relative abundance of each isoform and in the binding and activation capacities of the 14-3-3 isoforms.
Recent research has focused on one of the phosphorylation sites of AANAT located in the C-terminal region. We developed highly specific antisera to determine the phosphorylation state of the enzyme and revealed that the C-terminal site is phosphorylated at night and that the phosphorylation state of the enzyme decreases immediately after animals are exposed to light, most likely leading to destruction of the enzyme. We established that both the C- and N-terminal PKA sites are important for binding to 14-3-3 proteins. A remarkable on/off mechanism, centered around the C-terminal PKA site, switches the enzyme from highly active to highly inactive. In collaboration with P. Michael Iuvone, we found that the same mechanism operates in the retina to control both the activity of AANAT and melatonin production. In the retina, light acts directly on photoreceptor cells to control cyclic AMP, thereby controlling AANAT. We suspect that light acts through a similar mechanism in photosensitive pinealocytes. In collaboration with Philip Cole, Larry Szewcuk, Yousang Hwang, and Weiping Zhen, we used novel synthetic approaches to examine the role of phosphorylation in controlling AANAT. We made AANATs that contain non-hydrolyzable phosphate at the N-terminal PKA site, injected them into cells, and measured their biological half-life by using antisera. The results indicate that the non-hydrolyzable phosphate at the N-terminal PKA site prevents destruction of AANAT. We have also pursued new approaches to the development of AANAT inhibitors in order to identify lead compounds for drug development.
We established that, in the monkey pineal, large changes occur in the abundance of AANAT protein and activity, even though gene expression does not change, underscoring the importance of post-translational regulatory mechanisms in primate biology.
Molecular evolution of AANAT
We focused on an important element of the AANAT molecule—a floppy loop described as Loop 1, which forms a portion of the arylamine-binding pocket. Loop 1 has undergone notable changes during the course of the enzyme’s evolution. With Daniel Sackett and Sergio Hassan, we evaluated the importance of specific residues in the floppy loop to determine their contribution to the greater than 1,000-fold increase in specific activity of AANAT that occurred during chordate evolution, based on studies of Amphioxus, done by Pascaline Gaildrat and Jack Falcón. During the course of chordate evolution, Loop 1 acquired the tripeptide CPL65, which is responsible for increased enzyme activity. Removal of the CPL tripeptide markedly reduced activity. Moreover, increasing the local flexibility of the tripeptide region by P64G and P64A mutations had a counterintuitive effect, reducing both activity and the overall movement of Loop 1, as estimated from Langevin dynamics simulations. Binding studies indicate that the mutations increased the off-rate constant of a model substrate without altering the dissociation constant. The structural kink and local rigidity imposed by Pro-64 may enhance activity by favoring configurations of Loop 1 that facilitate catalysis without becoming immobilized by intramolecular interactions.
Studies conducted in collaboration with Jack Falcón revealed that there was a rapid expansion of the non-vertebrate type of AANAT in Amphioxus, resulting in seven copies. In addition, it was found that only the non-vertebrate form of the enzyme is present in this group. Studies on the eel and dogfish reveal the opposite—these chordates have only the vertebrate form of the enzyme. However, a striking exception was the discovery that the elephant fish, a member of the chimera sharks, has both vertebrate and non-vertebrate forms.
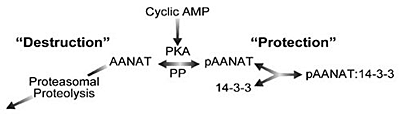
Figure 2.
Neuroepigenetics: global control of daily changes in pineal gene expression are regulated by an adrenergic/cyclic AMP mechanism
We initiated several projects aimed at (1) obtaining a global picture of differences in gene expression that occur on a night/day basis and (2) identifying genes that are highly enriched in the pineal gland. Beginnig with a relatively small number of genes, we can now, with microarrays, examine tens of thousands of genes and ESTs (expressed sequence tags). Together with Igor Dawid, we facilitated the NICHD-sponsored commercialization of an Affymetrix chip for analysis of gene expression in zebrafish and Xenopus. In collaboration with Dawid, Yoav Gothilf, and Reiko Toyama, we are characterizing pineal gene expression in the zebrafish and other species as a function of time of day and of development. We have already identified a set of genes highly expressed in the pineal gland. In collaboration with Peter Munson, David Carter, and Ruben Baler, we are analyzing the control of gene expression in second-messenger cascades. The work has triggered several investigations that analyze the expression of genes that previously had not appeared in the pineal literature, including those encoding methionine adenosyl transferase, MAP kinase phosphatase-1, PEPT1, phoshodiesterase 4D2, dopamine D4R receptor, and a subunit of the IgE receptor.
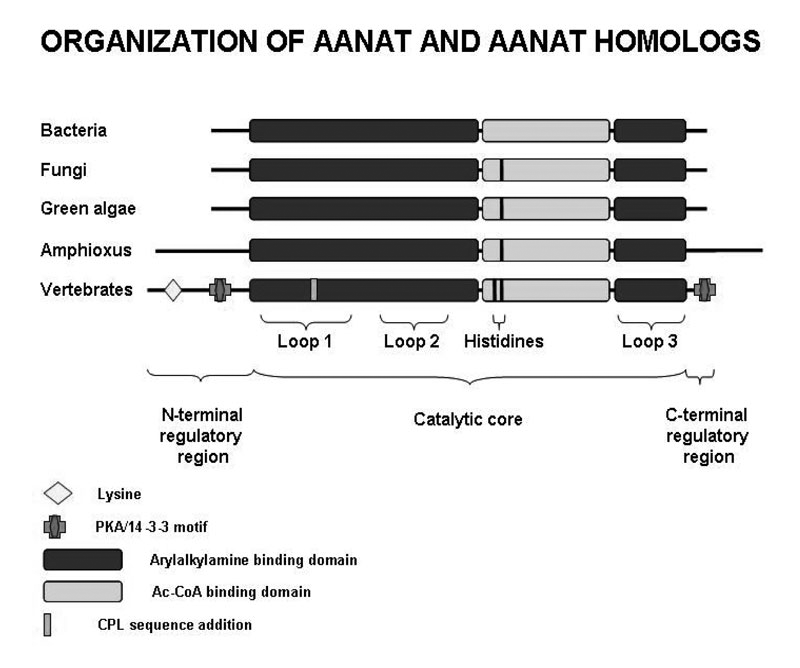
Click for a larger version.
Figure 3.
The group of pineal-specific genes expressed in the pineal gland of several vertebartes includes genes known to be associated with melatonin production and visual signal transduction. In addition, we have identified many genes (over 600), new to the pineal literature, that exhibit more than 2-fold night/day differences in expression and another set of genes that are exceptionally highly expressed in the pineal gland as compared with other tissues. This work is leading to a rapid increase in knowledge of the biochemical profile, conserved across species, of the pineal gland. It is also pointing to new transcriptional pathways controlled by previously unrecognized transcription factors; by analyzing these transcription factors and the promoters of genes that are either upregulated at night or highly expressed in the pineal gland, we will be able to construct a regulatory network that describes the cascade of transcription factors underlying the control of pineal gene expression. We have also identified potentially new and important pathways involved in cell-cell communication and signal transduction in the pineal gland; expression of some genes points to new functions for the pineal gland.
Using quantitative RT-PCR, our collaborator David Sugden has confirmed the results of our microarray studies and has provided a more detailed and quantitative description of the 24-hour patterns of gene expression, revealing that the patterns are not all similar or in phase but rather exhibit novel features. Together with our collaborator Morten Møller, using radiochemical in situ hybridization histology, we also confirmed the 24-hour patterns of rhythmic gene expression as well as the marked tissue specificity of genes of interest.
Our results permit a full-scale biochemical and physiological analysis of the control of genes in the pineal gland. Results from organ culture work revealed that the vast majority of genes that exhibit night/day changes in expression are also regulated by norepinephrine acting through a cyclic AMP mechanism that may involve cyclic AMP–response element binding sites in regulated genes and less specific epigenetic mechanisms.
Transcriptional control: NeuroD-1, CRX, OTX, Pax4, Pax6, and Mbnl2
Our microarray analyses, which were confirmed with independent techniques, revealed that the pineal gland expresses high levels of several transcription factors, including NeuroD1, CRX OTX, Pax4, and Pax6, that play a role in the development of the eye and other tissues. With collaborators under the direction of Morten Møller, we undertook a detailed description of the developmental appearance of these transcripts (Figure 4); the long-term plan is to determine the transcripts’ role in developmental and adult expression of genes in the pineal gland.
Otx2 is a vertebrate homeobox gene essential for the development of rostral brain regions. It also appears to play a role in the development of retinal photoreceptor cells and pinealocytes. We demonstrated the temporal expression pattern of Otx2 in the rat brain, with exceptionally strong expression in the pineal gland throughout late embryonic and postnatal stages. Widespread high expression of Otx2 in the embryonic brain becomes progressively restricted in the adult to the pineal gland. Crx (cone-rod homeobox), a downstream target gene of Otx2, showed a pineal expression pattern similar to that of Otx2, despite a distinct lag in time of onset. We identified the Otx2 protein in pineal extracts and found it to be localized in pinealocytes. Total pineal Otx2 mRNA neither showed night/day variation nor was influenced by the removal of sympathetic input, suggesting that the level of Otx2 mRNA is independent of the photoneural input to the gland. Our observations are consistent with the view that pineal expression of Otx2 is required for development and plays a role in the adult by controlling the expression of the cluster of genes associated with phototransduction and melatonin synthesis.
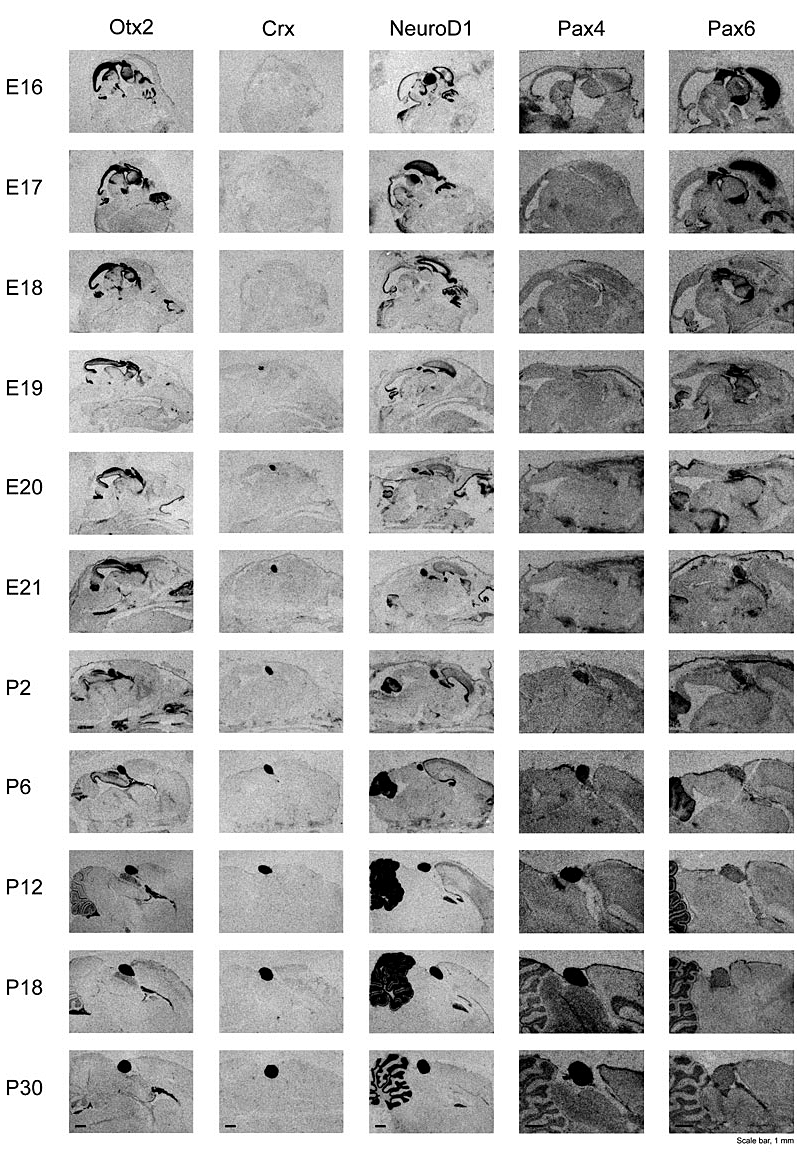
Click for a larger version.
Figure 4.
Developmental appearance of genes encoding the transcription factors Crx, Otx2, NeuroD1, Pax4, and Pax6 in the rat brain. The pineal gland is the heavily stained body most clearly visible in the Crx and Pax6 series. The method used to detect these transcripts is radiochemical in situ hybridization histology. These images are available in high resolution at science.nichd.nih.gov/confluence/display/sne/Developmental+Gallery.
NeuroD1/BETA2, a member of the bHLH transcription factor family, influences the fate of specific neuronal, endocrine, and retinal cells. In collaboration with Estela Muñoz, we found that NeuroD1 mRNA is highly abundant in the developing and adult rat pineal gland. Pineal expression begins in the 17-day embryo, at which time it is also detectable in other brain regions. Expression in the pineal gland increases during the embryonic period and is maintained thereafter at levels equivalent to those found in the cerebellum and retina. In contrast, NeuroD1 mRNA decreases markedly in non-cerebellar brain regions during development. Pineal NeuroD1 levels are similar during night and day and do not appear to be influenced by sympathetic neural input. Gene expression analysis of the pineal glands from neonatal NeuroD1 knockout mice identified 127 transcripts that are more than 2-fold downregulated and 16 that are more than 2-fold upregulated. Quantitative RT-PCR showed that the most dramatically downregulated gene is KIF5C (kinesin-family member 5C) (about 100-fold); the most dramatically upregulated gene is that encoding glutamic acid decarboxylase 1 (about 4-fold). Other affected transcripts encode proteins involved in differentiation, development, signal transduction, and trafficking. To elucidate the role of NeuroD1 in the rodent pinealocyte, we are using a Cre-Lox conditional knockout strategy to block expression of NeuroD1 in both the pineal gland and retina.
Our studies revealed that Pax4 is strongly expressed in the rat pineal gland and retina. The levels of pineal Pax4 transcripts are low in the fetus and increase postnatally; Pax6, on the other hand, exhibits an inverse pattern of expression and is more strongly expressed in the fetus. In the adult, the abundance of Pax4 mRNA exhibits a diurnal rhythm in the pineal gland, with maximal levels occurring late during the light period. Sympathetic denervation of the pineal gland by superior cervical ganglionectomy prevents the nocturnal decrease in pineal Pax4 mRNA. At night, the release of norepinephrine by sympathetic innervation adrenergically stimulates the pineal gland. We found that treatment with adrenergic agonists suppresses pineal Pax4 expression both in vivo and in vitro, suggesting that pineal Pax4 function is mediated by cyclic AMP, a second messenger of norepinephrine in the pineal gland. These findings suggest that the nocturnal decrease in pineal Pax4 mRNA is controlled by the sympathetic neural pathway, which, in turn, controls pineal function via an adrenergic-cyclic AMP mechanism. The daily changes in Pax4 expression may influence gene expression in the pineal gland.
Most recently, we have reported that the pineal gland expressed very high levels of muscleblind-like 2(Mbnl2), which encodes a zinc finger protein first identified in Drosophila. The gene appears to be essential for photoreceptor development and involved in RNA splicing. Mbnl2 is strongly expressed in the rat pineal gland. The abundance of pineal Mbnl2 transcripts follows a marked circadian rhythm with peak levels approximately sevenfold higher at night than day levels. Mbnl2 protein exhibits a similar rhythm. In vitro studies indicate that the abundance of Mbnl2 transcripts and protein are controlled by an adrenergic/cAMP mechanism.
Regulation of dopamine D4R receptor: cAMP + T3 "AND" gate regulation
Preliminary cDNA microarray studies suggested that D4R expression increases at night. Morten Møller confirmed such expression by in situ analysis, as we did by Northern blot and quantitative PCR. Other than trace levels, we could not find expression of other dopamine receptors in the pineal gland. D4R expression in the pineal gland is many-fold higher than in most other tissues, suggesting that the D4R receptor plays an important role in pineal signal transduction, perhaps as an inhibitor of adrenergic stimulation of adenylate cyclase. Regulation of the expression of this gene is unusual; in the case of other genes studied to date, including AANAT, MAT2, MKP-1, (Dio2, which encodes type II deiodinase), ICER, and FRA-2, the nocturnal increase in mRNA can be reproduced in organ culture by treatment with either norepinephrine or a cyclic AMP agonist. However, expression of D4R receptor mRNA does not increase following treatment with either norepinephrine or a cyclic AMP agonist. Rather, co-treatment with the thyroid hormone triiodothyronin (T3) and either norepinephrine or cyclic AMP is required to increase D4R mRNA, thereby establishing that expression of the D4R receptor is regulated by an “AND” gate operated by cAMP and T3 and providing the first evidence of a role for the thyroid gland in regulating a specific gene in the pineal gland. This finding is consistent with the knowledge of an increase in the capacity to convert thyroxin (T4) to T3 through induction of Dio2 at night in the pineal gland, indicating that melatonin synthesis might be a function of thyroid status. Our study thus has broad implications for dopamine signal transduction in other tissues.
As part of a broader effort to understand the similarities between the retina and pineal gland, a semiquantitative in situ hybridization study revealed prominent night/day variation in Drd4 expression in the retina of the rat, with a peak during the night. Drd4 expression is seen in all retinal layers, but the nocturnal increase is confined to the photoreceptors. Removal of sympathetic input to the eye does not affect retinal Drd4 expression, but T3 treatment induces Drd4 expression in the photoreceptors. In a developmental series, we found that expression of Drd4 is restricted to postnatal stages with a peak at postnatal day 12. The high Drd4 expression in the rat retinal photoreceptors during the night supports physiological and pharmacological evidence that the Drd4 receptor is involved in the dopaminergic inhibition of melatonin synthesis upon light stimulation. The sharp increase of Drd4 expression at a specific postnatal time suggests that dopamine is also involved in retinal development.
Role of islet antigen 2 and islet antigen 2beta in circadian biology
We discovered that the elimination of two proteins, islet antigen 2 and islet antigen 2beta, removes daily rhythms in activity, cardiovascular function, and temperature. The proteins are found in secretory vesicles and are best recognized by their association with type 1 diabetes; autoantibodies against the proteins are a predictor of the disease. Our studies suggest that the absence of daily rhythms may be attributable to a combination of effects on neurotransmission and downstream effects.
Additional Funding
- University of Copenhagen
Publications
- Rath M, Bailey MJ, Kim J-S, Ho AK, Gaildrat P, Coon SL, Møller M, Klein. Developmental and diurnal dynamics of Pax4 expression in the mammalian pineal gland: nocturnal down-regulation is mediated by adrenergic-cyclic adenosine 3’,5’-monophosphate signaling. Endocrinology 2009 150:803-811.
- Kim JS, Bailey MJ, Weller JL, Sugden D, Rath MF, Møller M, Klein DC. Thyroid hormone and adrenergic signaling interact to control pineal expression of the dopamine receptor D4 gene (Drd4). Mol Cell Endocrinol 2010 15:128-135.
- Kim JS, Coon SL, Weller JL, Blackshaw S, Rath MF, Møller M, Klein DC. Muscleblind-like 2: circadian expression in the mammalian pineal gland is controlled by an adrenergic-cAMP mechanism. J Neurochem 2009 110:756-764.
- Bailey MJ, Coon SL, Carter DA, Humphries A, Kim JS, Shi Q, Gaildrat P, Morin F, Ganguly S, Hogenesch JB, Weller JL, Rath MF, Møller M, Baler R, Sugden D, Rangel ZG, Munson PJ, Klein DC. Night/day changes in pineal expression of >600 genes: central role of adrenergic/cAMP signaling. J Biol Chem 2009 284:7606-7622.
- Kim SM, Power A, Brown TM, Constance CM, Coon SL, Nishimura T, Hirai H, Cai T, Eisner C, Weaver DR, Piggins HD, Klein DC, Schnermann J, Notkins AL. Deletion of the secretory vesicle proteins IA-2 and IA-2beta disrupts circadian rhythms of cardiovascular and physical activity. FASEB J 2009 23:3226-3232.
Collaborators
- Alastair Aitken, PhD, University of Edinburgh, Edinburgh, UK
- Michael J. Bailey, PhD, Texas A&M University, College Station, TX
- Ruben Baler, PhD, Laboratory of Cellular and Molecular Regulation, NIMH, Bethesda, MD
- Seth Blackshaw, PhD, The Johns Hopkins University, Baltimore, MD
- David Carter, PhD, University of Wales, Cardiff, UK
- Clivel Charlton, PhD, Florida A&M University, Tallahassee, FL
- Philippe Chemineau, PhD, Institut National de la Recherche Agronomique, Nouzilly, France
- Constance L. Chik, MD, University of Alberta, Edmonton, Canada
- Philip Cole, MD, The Johns Hopkins University, Baltimore, MD
- Cara M. Constance, PhD, College of the Holy Cross, Worcester, MA
- Igor Dawid, PhD, Program in Genomics of Differentiation, NICHD, Bethesda, MD
- Jack Falcón, PhD, Observeratoire Oceanologique/CNRS, Banyuls-sur-Mer, France
- Takaishi Furukawa, PhD, Osaka Bioscience Institute, Osaka, Japan
- Pascaline Gaildrat, PhD, Faculté de Médecin, Centre Hospitalier Universitaire, Rouen, France
- Vadivel Ganapathy, PhD, Medical College of Georgia, Augusta, GA
- Sandra Goebbels, PhD, Max-Planck-Institut für Experimentelle Medizin, Göttingen, Germany
- Yoav Gothilf, PhD, Tel Aviv University, Tel Aviv, Israel
- Sergio A. Hassan, PhD, Center for Molecular Modeling, CIT, NIH, Bethesda, MD
- Anthony Ho, PhD, University of Alberta, Edmonton, Canada
- John Hogenesch, PhD, Genome Institute of Novartis Foundation, San Diego, CA
- Ann Humphries, PhD, University of Wales, Cardiff, UK
- Yousang Hwang, PhD, The Johns Hopkins University, Baltimore, MD
- Ken-Ichi Inui, PhD, Kyoto University Hospital, Kyoto, Japan
- P. Michael Iuvone, PhD, Emory University School of Medicine, Atlanta, GA
- Lakshminarayan M Iyer, PhD, Computational Biology Branch, NCBL, Bethesda, MD
- Howard Jaffe, PhD, Laboratory of Neurochemistry, NINDS, Bethesda, MD
- Jong-So Kim, PhD, Pohang University of Science and Technology, Pohang, South Korea
- Ken Kirk, PhD, Laboratory of Chemistry, NIDDK, Bethesda, MD
- Eugene V. Koonin, PhD, National Center for Biotechnology Information, NLM, Bethesda, MD
- Benoit Malpaux, PhD, Institut National de la Recherche Agronomique, Nouzilly, France
- Stanford Markey, PhD, Laboratory of Neurotoxicology, NIMH, Bethesda, MD
- Morten Møller, PhD, Panum Institutet, Københavns Universitet, Copenhagen, Denmark
- Fabrice Morin, PhD, Université de Rouen, Mont-Saint-Aignan, France
- Estela Muñoz, PhD, Instituto de Histología y Embriología, Universidad Nacional de Cuyo, Mendoza, Argentina
- Peter Munson, PhD, Mathematical and Statistical Computing Laboratory, CIT, NIH, Bethesda, MD
- Abner Notkins, MD, Oral Infection and Immunity Branch, NIDCR, Bethesda, MD
- Tomas Obsil, PhD, Charles University, Prague, Czech Republic
- Hugh Piggins, PhD, Manchester University, Manchester, UK
- Zoila Rangel, MS, Mathematical and Statistical Computing Laboratory, CIT, NIH, Bethesda, MD
- Daniel Sackett, PhD, Program in Physical Biology, NICHD, Bethesda, MD
- Jürgen Schnermann, MD, Kidney Disease Branch, NIDDK, Bethesda, MD
- Peter J. Steinbach, PhD, Center for Molecular Modeling, CIT, NIH, Bethesda, MD
- David Sugden, PhD, Kings College, University of London, London, UK
- Larry Szewczuk, PhD, The Johns Hopkins University, Baltimore, Maryland
- Reiko Toyama, PhD, Program in Genomics of Differentiation, NICHD, Bethesda, MD
- David Weaver, PhD, University of Massachusetts, Worcester, MA
- Pin-Xian Xu, PhD, Mount Sinai School of Medicine, New York, NY
- Al Yergey, PhD, Mass Spectrometry Core Facility, NICHD, Bethesda, MD
- Weiping Zhen, PhD, The Johns Hopkins University, Baltimore, MD
Contact
For more information, email kleind@mail.nih.gov or visit sne.nichd.nih.gov.