

You are here: Home > Section on Cellular Differentiation
Molecular Genetics of Heritable Human Disorders

- Janice Y. Chou, PhD, Head, Section on Cellular Differentiation
- Hyun-Sik Jun, PhD, Staff Scientist
- Young Mok Lee, PhD, Visiting Fellow
- Su Ru Lin, PhD, Visiting Fellow
- Brian C. Mansfield, PhD, Guest Researcher
- Chi-Jiunn Pann, BS, Senior Research Assistant
- Yong Jun Lee, PhD, Visiting Fellow
- Lane H. Wilson, BS, Postbaccalaureate Fellow
We conduct research both to delineate the pathophysiology of glycogen storage disease type I (GSD-I) and glucose-6-phosphatase-beta (G6Pase-beta or G6PC3) deficiency and to develop novel therapies for these disorders. There are two types of GSD-I: GSD-Ia, a deficiency in G6Pase-alpha (or G6PC); and GSD-Ib, a deficiency in the glucose-6-phosphate transporter (G6PT). A third disease, G6Pase-beta deficiency, also known as severe congenital neutropenia syndrome type 4 (SCN4), is grouped with GSD-I. G6Pase-alpha and G6Pase-beta are endoplasmic reticulum (ER)–bound glucose-6-phosphate (G6P) hydrolases, with active sites inside the lumen; the hydrolases depend on G6PT to translocate G6P from the cytoplasm into the ER lumen. The G6PT/G6Pase-alpha complex maintains interprandial glucose homeostasis while the G6PT/G6Pase-beta complex maintains neutrophil energy homeostasis and functionality. GSD-Ia and GSD-Ib patients manifest a common metabolic phenotype of impaired glucose homeostasis not shared by G6Pase-beta deficiency. GSD-Ib and G6Pase-beta–deficient patients manifest a common myeloid phenotype of neutropenia and neutrophil dysfunction not shared by GSD-Ia. Neutrophils express the G6PT/G6Pase-beta complex, and inactivation of G6PT or G6Pase-beta leads to enhanced neutrophil apoptosis, which underlies neutropenia in GSD-Ib and G6Pase-beta deficiency. More recently, we showed that G6Pase-beta is essential for neutrophil energy homeostasis. G6Pase-beta deficiency prevents recycling of ER glucose to the cytoplasm, leading to neutrophil dysfunction. There is no cure for GSD-I and G6Pase-beta deficiency. Animal models of the three disorders are available and are being exploited to both delineate the disease more precisely and develop new treatment approaches, including gene therapy.
Lack of glucose recycling between the ER and cytoplasm underlies neutrophil dysfunction in G6P-ase deficiency.
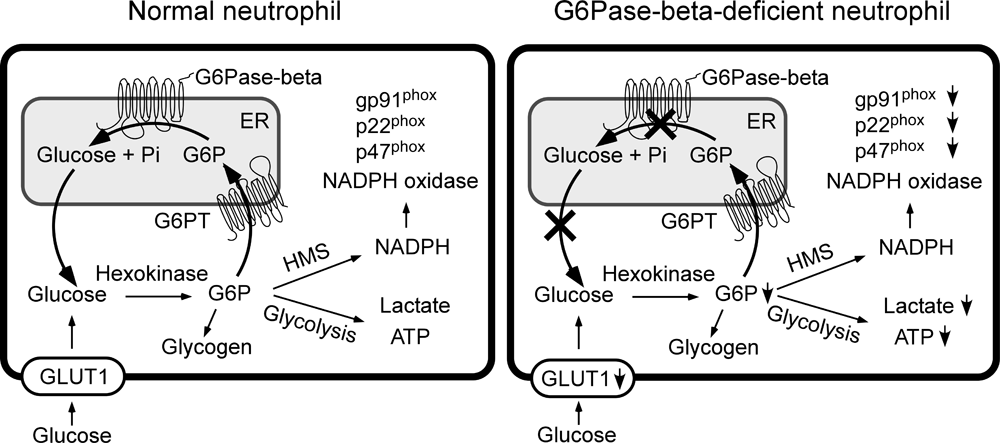
Click image to enlarge.
Figure 1. Proposed pathways for G6P metabolism in normal and G6Pase-beta−deficient neutrophils
Glucose transported into the cytoplasm via GLUT1 is metabolized by hexokinase to G6P, which can participate in glycolysis, hexose monophosphate shunt (HMS), and glycogen synthesis or be translocated into the lumen of the ER by the G6PT. In normal neutrophils, G6P localized within the ER lumen can be hydrolyzed by G6Pase-beta, with the resulting glucose transported back into the cytoplasm to re-enter any of the previously mentioned cytoplasmic pathways. However, in G6Pase-beta–deficient neutrophils, which lack a functional G6Pase-beta, ER−localized G6P cannot be recycled to the cytoplasm. The GLUT1 transporter, responsible for the transport of glucose into and out of the cell, is shown embedded in the plasma membrane. Both the G6PT transporter, which is responsible for the transport of G6P into the ER, and G6Pase-beta, which is responsible for hydrolyzing G6P to glucose and phosphate, are shown embedded in the ER membrane.
G6Pase-beta (or G6PC3) deficiency, characterized by neutropenia and neutrophil dysfunction, is caused by deficiencies in G6Pase-beta, which converts G6P into glucose, the primary energy source of neutrophils. Enhanced neutrophil ER stress and apoptosis underlie neutropenia in G6Pase-beta deficiency, but the exact functional role of G6Pase-beta in neutrophils remains unknown. We hypothesized that the ER recycles G6Pase-beta–generated glucose to the cytoplasm, thus regulating the amount of available cytoplasmic glucose/G6P in neutrophils. Accordingly, a G6Pase-beta deficiency would impair glycolysis and hexose monophosphate shunt activities, leading to reductions in lactate production, ATP production, and NADPH oxidase activity (Figure 1). Using non-apoptotic neutrophils, we showed that glucose transporter-1 (GLUT1) translocation is impaired in neutrophils from G6Pase-beta–deficient patients and mice along with impaired glucose uptake in murine G6Pase-beta–deficient neutrophils (Figure 1). Moreover, levels of G6P, lactate, and ATP are markedly lower in human and murine G6Pase-beta–deficient neutrophils than in their respective controls. In parallel, the expression of NADPH oxidase subunits and membrane translocation of p47phox are down-regulated in human and murine G6Pase-beta–deficient neutrophils (Figure 1). The results establish that, in neutrophils, G6Pase-beta is essential for normal energy homeostasis. G6Pase-beta deficiency prevents recycling of ER glucose to the cytoplasm, leading to neutrophil dysfunction.
G-CSF improves G6Pase-beta−deficient neutrophil function by modulating apoptosis and energy homeostasis
G6Pase-beta deficiency underlies a congenital neutropenia syndrome in which neutrophils exhibit enhanced ER stress, elevated apoptosis, impaired energy homeostasis, and impaired functionality. We showed that murine G6Pase-beta–deficient neutrophils undergoing ER stress activate protein kinase–like ER kinase and phosphatidylinositol 3,4,5-trisphosphate (PtdIns(3,4,5)P3)/Akt signaling pathways and that neutrophil apoptosis is mediated, in part, by the intrinsic mitochondrial pathway. In G6Pase-beta–deficient patients, granulocyte colony stimulating factor (G-CSF) improves neutropenia, but its impact on neutrophil apoptosis and dysfunction is unknown. We showed that G-CSF delays neutrophil apoptosis in vitro by modulating apoptotic mediators. However, in the presence or absence of G-CSF, G6Pase-beta–deficient neutrophils in culture exhibit accelerated apoptosis compared with wild-type neutrophils. Limiting glucose (0.6 mM) accelerates apoptosis but is more pronounced for wild-type neutrophils, leading to similar survival profiles for both neutrophil populations. In vivo G-CSF therapy completely corrects neutropenia and normalizes levels of p-Akt, PtdIns(3,4,5)P3, and active caspase-3. Neutrophils from in vivo G-CSF−treated G6Pase-beta−deficient mice exhibit increased glucose uptake and elevated intracellular levels of G6P, lactate, and ATP, leading to improved functionality. Together, the results strongly suggest that G-CSF improves murine G6Pase-beta−deficient neutrophil survival by modulating apoptotic mediators and rectifies function by enhancing energy homeostasis.
Complete normalization of hepatic G6Pase-alpha deficiency in murine GSD-Ia using gene therapy
GSD-Ia patients deficient in G6Pase-alpha (or G6PC) manifest impaired glucose homeostasis. We examined the efficacy of liver G6Pase-alpha delivery mediated by AAV-GPE, an AAV serotype 8 vector expressing human G6Pase-alpha directed by the human G6PC promoter/enhancer (GPE), and compared it with AAV-CBA, which directed murine G6Pase-alpha expression, by using a hybrid chicken beta-actin (CBA) promoter/CMV enhancer. AAV-GPE–directed hepatic G6Pase-alpha expression in the infused G6Pase-alpha–null mice declined 12-fold from age 2 to 6 weeks but stabilized at wild-type levels from age 6 to 24 weeks. In contrast, the expression directed by AAV-CBA declined 95-fold over 24 weeks, demonstrating that the GPE is more effective in directing persistent in vivo hepatic transgene expression. We further showed that the rapid decline in transgene expression directed by AAV-CBA results from an inflammatory immune response elicited by the AAV-CBA vector. The AAV-GPE–treated G6Pase-alpha–null mice exhibit normal levels of blood glucose, blood metabolites, hepatic glycogen, and hepatic fat. Moreover, the mice maintained normal blood glucose levels even after six hours of fasting. The complete normalization of hepatic G6Pase-alpha deficiency by the G6PC promoter/enhancer holds promise for the future of gene therapy in human GSD-Ia patients.
SLC37A1 and SLC37A2 are phosphate-linked, G6P antiporters.
Blood glucose homeostasis between meals depends on production of glucose within the ER of the liver and kidney by hydrolysis of G6P into glucose and phosphate (Pi). This reaction depends on coupling the G6PT with G6Pase-alpha. G6PT belongs to the SLC37 family of membrane-bound transporters. Only G6PT, also known as SLC37A4, has been characterized, and it acts as a Pi-linked G6P antiporter. The other three SLC37 family members, predicted to be sugar-phosphate:Pi exchangers, have not been characterized functionally. Using reconstituted proteoliposomes, we examined the antiporter activity of the other SLC37 members along with their ability to couple with G6Pase-alpha. We showed that SLC37A1 and SLC37A2 are ER-associated, Pi-linked antiporters that can transport G6P. The activity of SLC37A3 is unknown. Unlike G6PT, neither SLC37A1 nor SLC37A2 is sensitive to chlorogenic acid, a competitive inhibitor of physiological ER G6P transport, and neither couples to G6Pase-alpha. We concluded that three of the four SLC37 family members are functional sugar-phosphate antiporters. However, only G6PT/SLC37A4 matches the characteristics of the physiological ER G6P transporter, suggesting the other SLC37 proteins play roles independent of blood glucose homeostasis.
Additional Funding
- The Children's Fund for Glycogen Storage Disease Research
Publications
- Chou JY, Jun HS, Mansfield BC. Glycogen storage disease type I and G6Pase-β deficiency: etiology and therapy. Nat Rev Endocrinol 2010;6:676-688.
- Yiu WH, Lee YM, Peng WT, Pan C-J, Mead PA, Mansfield BC, Chou JY. Complete normalization of hepatic G6PC deficiency in murine glycogen storage disease type Ia using gene therapy. Mol Ther 2010;18:1076-1084.
- Jun HS, Lee YM, McDermott DH, DeRavin SS, Murphy PM, Mansfield BC, Chou JY. Lack of glucose recycling between endoplasmic reticulum and cytoplasm underlies cellular dysfunction in glucose-6-phosphatase-beta-deficient neutrophils in a congenital neutropenia syndrome. Blood 2010;116:2783-2792.
- Jun HS, Lee YM, Song KD, Mansfield BC, Chou JY. G-CSF improves murine G6PC3-deficient neutrophil function by modulating apoptosis and energy homeostasis. Blood 2011;117:3881-3892.
- Pan CJ, Chen SY, Jun HS, Lin SR, Mansfield BC, Chou JY. SLC37A1 and SLC37A2 are phosphate-linked, glucose-6-phosphate antiporters. PloS ONE 2011;in press.
Collaborators
- Reba Condiotti, PhD, Hadassah University Hospital, Jerusalem, Israel
- Suk See DeRavin, MD, Laboratory of Host Defenses, NIAID, Bethesda, MD
- Alessandra Eva, PhD, Gaslini Institute, Genova, Italy
- David H. McDermott, MD, Laboratory of Host Defenses, NIAID, Bethesda, MD
- Philip M. Murphy, MD, Laboratory of Host Defenses, NIAID, Bethesda, MD
- Luigi Varesio, PhD, Gaslini Institute, Genova, Italy
- David A. Weinstein, MD, University of Florida College of Medicine, Gainesville, FL
Contact
For further information, contact chouja@mail.nih.gov.