

You are here: Home > Section on Genetic Disorders of Drug Metabolism
Molecular Biology, Regulation, and Biochemistry of UDP–Glucuronosyltransferase Isozymes

- Ida S. Owens, PhD, Head, Section on Genetic Disorders of Drug Metabolism
- Nikhil K. Basu, PhD, Staff Scientist
- Amit Raychoudhuri, PhD, Visiting Fellow
- Mousumi Basu, BS, Special Volunteer
UDP–glucuronosyltransferase (UGT) isozymes–distributed primarily in liver, kidney, the gastrointestinal tract, and steroid-responsive tissues–are known to carry out the essential function of converting innumerable structurally diverse lipophilic endogenous substrates, such as neurotoxic bilirubin and catechol estrogens and dihydrotestosterone, and dietary aromatic-like therapeutics to water-soluble excretable glucuronides. Most importantly, environmental pro-carcinogens and contaminants derived from pyrolysates are converted to avoid chemical toxicities. Our studies demonstrated that each UGT isozyme so far examined requires on-going regulated phosphate signaling, which enables an active site to convert an unspecified number of substrates. Recently, further studies showed that the human prostate luminal-cell UGT-2B15 and basal-cell UGT-2B17, which are 97% identical, have an additional Src or Src/PKCε–partnership phosphorylation site, respectively, at position 98–100. We found that the two isozymes exhibit opposite behavior when their Src sites are compromised: UGT-2B15 becomes polyubiquitinated, thus exhibiting a pro-apoptotic effect, while the activity of UGT-2B17 is elevated by 50%. Our studies will thus continue to detail and understand the specific reactions involved in human prostate–luminal cell apoptosis and de-ubiquitination. In collaboration with ongoing research within the NCI, NIDCR, and with researchers at the University of Maryland, we will also carry out basic studies to better understand prostate cancer development.
Two dihydrotestosterone-metabolizing UDP–glucuronosyltransferases control pro-apoptosis and anti-apoptosis in preventing prostate transformation and its continuity and may thus serve as clinical markers.
Click image to enlarge.
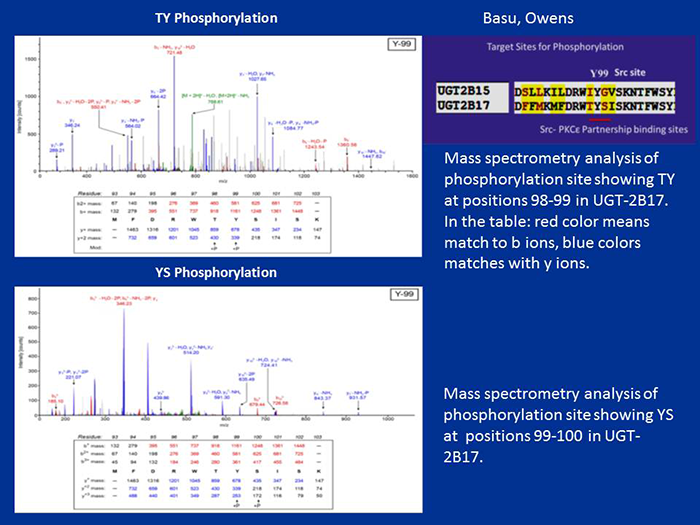
Figure 1. Mass-spectrometric analysis of TpYpSp phosphorylation of UGT-2B17
Phosphorylation of UGT-2B17 at 98–100 (TYS) residues
COS-1 cell–expressed human UGT-2B17 metabolizes primarily dihydrotestosterone (DHT) and its androstanediol (ADT-diol) metabolite. Reversible or irreversible inhibition following treatment with curcumin or general PKC inhibitors, respectively, is evidence that the prostate-distributed isozyme requires regulated phosphorylation. Treatment of COS 1–expressed UGT-2B17 with PKCalpha-siRNA (small interfering RNA) downregulates activity while treatment with PKCepsilon-siRNA, Src-siRNA, or an Src kinase–specific inhibitor (PP2) upregulates UGT-2B17's activity, indicating that PKCalpha activates UGT-2B17 whereas PKCepsilon and/or Src downregulate its activity. The PKC-site mutant S172A-UGT-2B17 is a null mutant while the S422A-UGT-2B17 mutant is unchanged. The tyrosine kinase–site mutants Y99F-UGT-2B17 and Y237F-UGT-2B17 show preferential inhibition of DHT over ADT-diol glucuronidation at the 70/30% and 90/60% levels, respectively. When expressed in Src–free cells, the respective inhibition is 50% to 60/70%. Solubilized UGT-2B17–His—transfected into Src–free fibroblasts and subjected to in vitro 33P-ATP–dependent phosphorylation by PKCepsilon and/or Src—revealed higher radiolabeling of 55- to 58-kDa UGT-2B17-His than phosphorylation by PKCalpha and/or Src. The findings indicate that UGT-2B17 requires regulated phosphorylation by PKCalpha, PKCepsilon, and Src, which is consistent with the complexity of synthesis and metabolism of its major substrate, DHT.
Click image to enlarge.
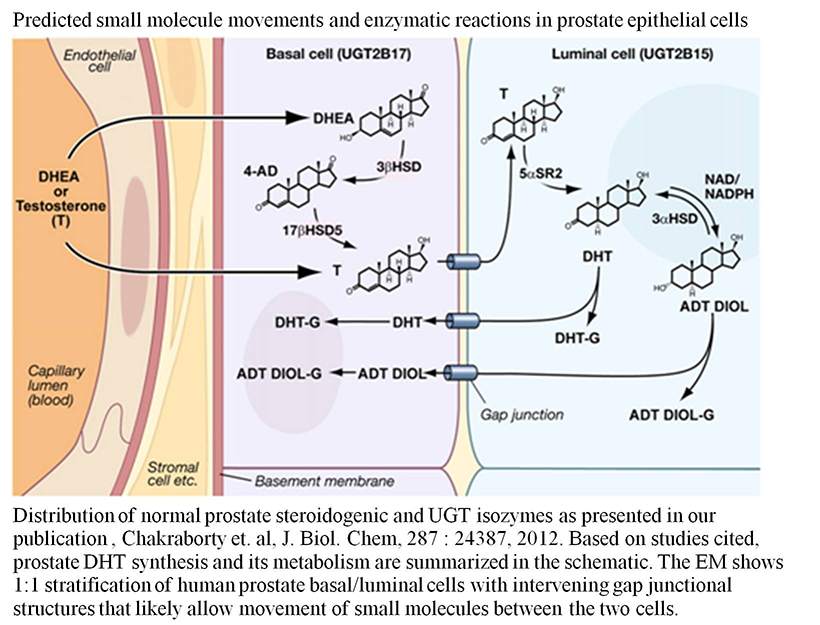
Figure 2. Predicted small-molecule movements and enzymatic reactions in prostate epithelial cells
Image taken from S. K. Chakraborty et al., J Biol Chem 2012;287:24387.
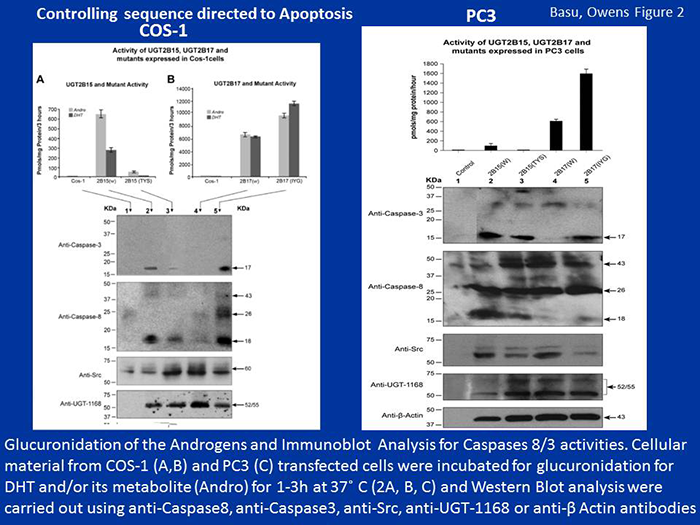
Human prostate basal cell–distributed UGT-2B17 and its UGT-2B15 homolog, which are 97% identical, metabolize both DHT and its ADT-diol metabolite. Mass spectrometry confirmed that UGT-2B17 has 4 of the 5 predicted phosphorylation sites found in luminal cell–distributed UGT-2B15. However, unlike UGT-2B15, UGT-2B17 contains the rare tri-phosphorylated TYS sequence at position 98–100. Both isozymes undergo regulated phosphorylation for two different functions. Based on the analysis of 6/19 human UGTs, using anti-PKCepsilon immunocomplexes of UGT-2B17, we found that the Src/PKCepsilon phosphorylation site TYS plays a critical role. Following UGT-2B17 expression in Src−/− or Src+/− cells, glucuronidation of DHT or its ADT-diol metabolite indicates that Src inhibits both by 50%, which necessarily elevates anti-apoptotic DHT levels. Switching the IYG of wild-type UGT-2B15(IYG) for the TYS in wild-type UGT-2B17(TYS) at positions 98–100, followed by expression in COS-1 or PC3 cells (human prostate cancer cell lines), enabled mutant UGT-2B17(IYG) to activate caspases 8/3 (apoptotic enzymes) 4- to 10-fold more than wild-type UGT-2B15(IYG), while mutant UGT-2B15(TYS) suppressed activation of caspases 8/3 by over 50% of wild-type UGT-2B15 (IYG) levels. Treatment of LNCaP cells (androgen-sensitive prostate adenocarcinoma cells), which contain endogenous UGT-2B15 and UGT-2B17, with the Src inhibitor curcumin enabled a cycle of apoptosis and recovery within 16 h. Combined, the evidence indicates that the triple-phosphorylated TYS in UGT-2B17 creates a signaling site involving Src and PKCepsilon that is anti-apoptotic, while the pro-apoptotic Src–specific binding/phosphorylation site IYG in UGT-2B15, at position 98–100, is blocked by Src.
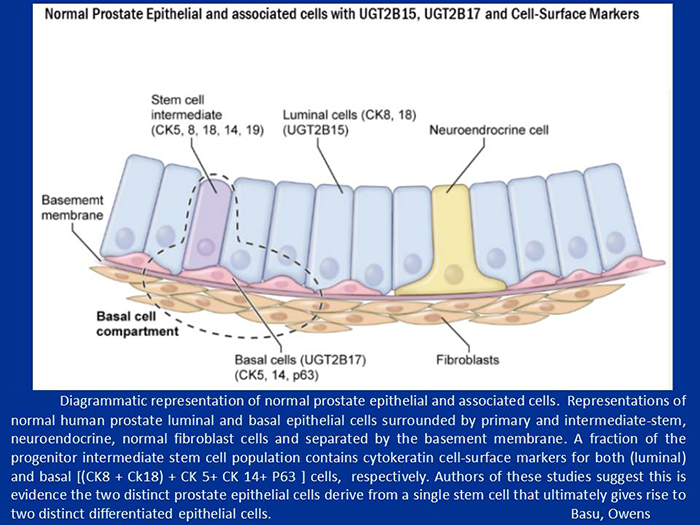
We have thus shown that the 97% identical prostate basal cell–distributed UGT2B17 and luminal cell–distributed UGT2B15 are programmed by phosphorylation to act in an opposite fashion (Basu NK et al., under revision). Further, we have provided evidence that human prostate luminal cell–distributed DHT–metabolizing UGT-2B15 uses programmed phosphorylation–based signaling to carry out luminal cell–specific apoptosis, whereas basal cell–distributed UGT-2B17—contained within a basal-cell compartment that houses intermediate stem cells with both basal and luminal cell–surface cytokeratin markers—utilizes programmed anti-apoptotic signaling to protect this population of cells. For the first time, we identified the luminal cell–specific UGT-2B15 as the pro-apoptotic agent, which removes challenged luminal cells before transformation can ensue. Moreover, we identified basal cell–specific UGT-2B17 as the anti-apoptotic agent that protects intermediate stem cells containing both basal and luminal cell–surface markers, which replace the departed luminal cell and give rise to a new basal cell.
In summary, our findings indicate that UGT-2B15 controls luminal transformation and that UGT-2B17 controls continuity of the prostate. Endogenously expressed UGT-2B15/UGT-2B17 in LNCaP cells show a cycle of apoptosis and recovery within 16 h after treatment with the Src inhibitor curcumin, and we found evidence that the triple-phosphorylated TYS creates an anti-apoptotic Src/PKCepsilon signaling site, while the pro-apoptotic Src–specific IYG signaling site in UGT-2B15 is blocked by Src phosphorylation. Hence, UGT-2B15 and UGT-2B17 appear to be unique clinical markers (Owens et al., Proc Natl Acad Sci USA 2014;submitted).
Click image to enlarge.
Figure 3. Controlling sequences for apoptosis
Controlling sequences for pro-apoptosis and anti-apoptosis in UGT-2B15 and UGT-2B17 expressed in COS-1 cells and the aggressive PC3 prostate cell line
Prostate-distributed mouse Ugt2b34 and Ugt2b36 control estrogenic metabolites.
To establish an in vivo mammary-gland model that prevents depurination by 4-OH-catecholestrogens associated with the initiation of carcinogenesis, we pursued studies to identify mouse homologs of the highly effective human UGT-2B7. Using sequence analysis, we found that mouse Ugt-2b34 and Ugt-2b36 homologs avidly metabolize the test agent 4-hydroxyestrone, with Ugt-2b35 expressing trivial activity. Unlike low Km UGT-2B7 (14M), Ugt-2b34 and Ugt-2b36 metabolized 4-hydroxyestrone with 90M Km and 430M Km, respectively. Unexpectedly, the mouse isozymes are distributed primarily in male hormone–responsive tissues, whereas human UGT-2B7 is found primarily in female hormone–responsive tissues. Also, we found that Ugt-2b34 metabolizes the non-classical estrogenic DHT metabolite ADT-diol at a greater rate than DHT, which is not known to be estrogenic. Notably, UGT-2B7 does not metabolize xeno-estrogens; Ugt-2b34 and Ugt-2b36 did, however, metabolize bisphenol A (BPA) and diethylstilbestrol (DES) at superior rates. We also found, through real-time PCR–based analysis of estrogen receptor alpha (Esr1) gene knockout in mouse prostate, 50% and 63% lower Ugt2b34 mRNA and Ugt2b36 mRNA levels, respectively, than in controls. However, estrogen receptor beta (Esr2) knockout (KO) revealed a 2.7/3.3-fold increase in Ugt-2b34 mRNA and Ugt-2b36 mRNA, respectively, in prostate. Esr1 KO completely suppressed Ugt-2b34 and Ugt-2b36 mammary-gland mRNA; Esr2 KO caused a 12-fold increase in Ugt-2b34 mRNA without affecting Ugt-2b36 mRNA. Hence, according to tissue-distribution studies it appears that male mice benefit from both Ugt isoforms, while females benefit from only one Ugt. Our findings for Ugt-2b34 and Ugt-2b36 suggest that the two mouse isozymes are intrinsically programmed to protect against a more complex environment than are human high-activity UGT-2B7 and low-activity UGT-2B4 isozymes (Raychoudhuri A et al., Biosci Rep 2014;in press).
Click image to enlarge.
Figure 4. Diagrammatic representation of normal prostate epithelial and associated cells
Normal prostate epithelial and associated cells with UGT-2B15, UGT-2B17, and cell surface markers.
Publications
- Basu NK, Kole L, Basu M, Chakraborty K, Mitra PS, Owens IS. The major chemical detoxifying system of UDP-glucuronosyltransferases requires regulated phosphorylation supported by protein kinase C. J Biol Chem 2008;283:23048-23061.
- Banerjee R, Pennington MW, Garza A, Owens IS. Mapping the UDP glucuronic acid binding site in UDP-glucuronosyltransferase-1A10 by homology-based modeling: confirmation with biochemical evidence. Biochemistry 2008;47:7385-7392.
- Mitra PS, Basu NK, Owens IS. Src supports UDP-glucuronosyltransferase-2B7 detoxification of catechol estrogens associated with breast cancer. Biochem Biophys Res Commun 2009;382:651-656.
- Mitra PS, Basu NK, Basu M, Chakraborty S, Saha T, Owens IS. Regulated phosphorylation of a major UDP-glucuronosyltransferase isozyme by tyrosine kinases dictates endogenous substrate selection for detoxification. J Biol Chem 2011;286:1639-1648.
- Chakraborty SK, Basu NK, Jana S, Basu M, Raychoudhuri A, Owens IS. Protein kinase Calpha and Src kinase support human prostate-distributed dihydrotestosterone-metabolizing UDP-glucuronosyltransferase 2B15 activity. J Biol Chem 2012;287:24387-24396.
Collaborators
- Praveen Arany, BDS, MDS, MMSc, PhD, Oral and Pharyngeal Cancer Branch, NIDCR, Bethesda, MD
- James L. Gulley, MD, PhD, FACP, Genitourinary Malignancies Branch, Center for Cancer Research, NCI, Bethesda, MD
- Antony McDonagh, PhD, University of California San Francisco, San Francisco, CA
- Zhihong Nie, PhD, Maryland Nanocenter, University of Maryland, College Park, MD
- Juan Rivera, PhD, Molecular Immunology and Inflammation Branch, NIAMS, Bethesda, MD
Contact
For further information, contact ida.owens2@nih.gov.