

You are here: Home > Section on Molecular Endocrinology
Receptors and Actions of Peptide Hormones and Regulatory Proteins in Endocrine Mechanisms

- Maria L. Dufau, MD, PhD, Head, Section on Molecular Endocrinology
- Chon-Hwa Tsai-Morris, PhD, Staff Scientist
- Raghuveer Kavarthapu, PhD, Postdoctoral Fellow
- Peng Zhao, PhD, Postdoctoral Fellow
- Ruifeng Yang, MSc, Predoctoral Fellow
We investigate the molecular basis of peptide hormone control of gonadal function, with particular emphasis on the structure and regulation of the genes encoding the luteinizing hormone receptor (LHR) and prolactin (PRL) receptor (PRLR). We also investigate the regulatory mechanism(s) involved in the progress of spermatogenesis and the control of Leydig cell (LC) function. Our studies focus on the regulation of human LHR transcription (nuclear orphan receptors, epigenetics, DNA methylation, second messengers, repressors, corepressors, and coactivators), as well as on the multiple-promoter control of hPRLR gene transcription. We are elucidating the functions of two inhibitory short forms of prolactin receptors and their impact on the long form of the receptor as well as their relevance to physiological regulation and breast cancer. We also investigate novel gonadotropin-regulated genes of relevance to the progression of testicular gametogenesis, LC function, and other endocrine processes. We focus on the function and regulation of the gonadotropin-regulated testicular RNA helicase (GRTH/DDX25), an essential post-transcriptional regulator of spermatogenesis that was discovered, cloned, and characterized in our laboratory. The various functions of GRTH/DDX25 provide a fertile ground for the development of a male contraceptive.
The luteinizing hormone receptor
The luteinizing hormone receptor (LHR) is expressed primarily in the gonads, where it mediates luteinizing hormone (LH) signals that regulate cyclic ovarian changes or testicular function. Transcription of the LHR gene is regulated by complex and diverse networks, in which coordination and interactions between regulatory effectors are essential for silencing/activation of LHR expression. The proximal Sp1 site of the promoter recruits histone (H) deacetylases and the Sin3A corepressor complex that contributes to the silencing of LHR transcription. Site-specific acetylation/methylation-induced phosphatase release serves as an on switch for Sp1 phosphorylation at Ser641, which causes p107 repressor release from Sp1, recruitment of TFIIB and Pol II, and transcriptional activation. Maximal derepression of the gene is dependent on DNA methylation of the promoter, H3/H4 acetylation, and HDAC/Sin3 A release. Positive Cofactor 4 (PC4) has an important role in the formation assembly of the preinitiation complex (PIC) in trichostatin A (TSA)–mediated LHR transcription (1). It is recruited by Sp1 following TSA treatment and acts as its coactivator. However, PC4 does not participate in TSA release of phosphatases, Sp1 phosphorylation, or release of repressor/ complexes. Although TFIIB recruitment is dependent on PC4, we ruled out TFIIB as its direct target and acetylation of PC4 in the activation process. However, we demonstrated TSA–induced acetylation of PC4–interacting proteins, identified as acetylated H3, and we demonstrated, by ChIP/reChIP, PC4's presence in the complex in association with chromatin at the promoter. We are now proceeding to determine the sites of acetylation at H3. The role of PC4–acetylated H3 interactions in chromatin structure and their participation in the assembly of the PIC and transcriptional activation are under investigation.
Gonadotropin-regulated testicular RNA helicase
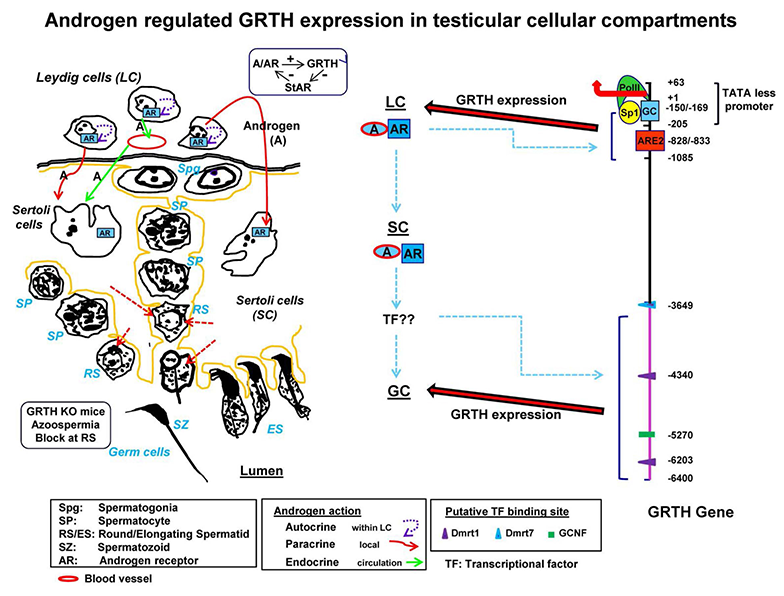
Click image to enlarge.
Figure 1. Diagram of 5′ flanking sequence of the GRTH gene that directs its cell-specific expression in testicular cells and direct/indirect actions of androgen on GRTH transcription in cellular compartments.
Androgen (A) produced from Leydig cells (LC) binds to androgen receptor (AR) and subsequently interacts with androgen response element (ARE2) (−828/−833) to direct GRTH gene expression in LC (3). Paracrine activation by androgen from LC induces A/AR–responsive gene(s) expression in Sertoli cells such as th transcription factor (TF) reproductive homeobox on chromosome X5 or others to be identified. In turn, these may activate downstream targets that directly or indirectly induce GRTH transcription through TF binding to 5′ elements located in the distal (−3600/−6450 bp) 5′ flanking region of the gene (i.e., Dmrt1/7, GCNF/RTR or others) (4).
Gonadotropin-regulated testicular RNA helicase (GRTH, also known as DDX25) is a testis-specific member of the DEAD-box family of RNA helicases present in LCs and meiotic germ cells. The multi-functional protein is essential for the completion of spermatogenesis. Males lacking GRTH are sterile owing to the absence of sperm resulting from failure of round spermatids to elongate. In addition, to its intrinsic RNA helicase activity, GRTH is a shuttling protein that exports specific mRNAs from the nucleus to cytoplasmic sites. Our studies demonstrated the essential participation of the GRTH export/transport of mRNAs in the structural integrity of the Chromatoid Body (storage/processing of mRNAs) and their transit/association to actively translating polyribosomes, where GRTH may regulate translational initiation of genes. We identified mRNAs that are associated with GRTH and regulated at polysomal sites of cell populations of the mouse testis (2). The reduction in mRNAs associated at polysomal sites in the differential studies (knockout compared with wild type), which is not detected at the total cellular level but in the cytoplasm with abolition of protein expression, reflects the importance of the transport function of GRTH to relevant sites and underscore its impact in protein synthesis.
GRTH is regulated by LH through androgen (A) at the transcriptional level in LCs (direct) and germ cells (presumably indirect) of the testis, where its expression is both cell- and stage-specific. The helicase displays a novel negative autocrine control of A production in LCs by preventing overstimulation of the LH–induced A pathway through enhanced degradation of the StAR protein. Our studies described the mechanism by which A/androgen receptor (AR) regulates the expression of the GRTH gene in the LC via a short-range chromosomal loop (3). Through its activation of GRTH transcription, A/AR signaling in LCs participates in an autocrine regulatory mechanism with a major impact on LC steroidogenic function.
Our development of transgenic mice model carrying GRTH 5′ flanking region–GFP reporter provides a unique in vivo system that permits differential elucidation of regulatory regions upstream in the GRTH gene that direct the gene's expression in germ cells (pachytene spermatocytes and round spermatids) and downstream in LCs (4). We identified binding sites for germ cell–specific transcription factors (GCNF, Dmrt1/7) in the distal region. In contrast, the proximal region directs basal GRTH expression and A-induced intracrine expression in LCs through a functional androgen-response element (ARE). In the transgenic animal model, the AR antagonist flutamide blocked GRTH–GFP expression in LCs and germ cells, demonstrating direct (intracrine regulation by A/AR in LCs) and indirect effects of A/AR in germ cells, which do not express AR, through paracrine regulation by A/AR in Sertoli cells. The model permits us to elucidate the mechanism of androgen action in germ cells and thus would permit the identification of androgen-regulated factors that control expression of a critical gene(s) required for GRTH expression in germ cells involved in the progress of spermatogenesis. This could lead to development of contraceptive strategies in Sertoli cells to block sperm formation without impacting other aspects of androgen action.
The prolactin receptor
The PRL receptor (PRLR) is a member of the lactogen/cytokine receptor family, which mediates the diverse cellular actions of PRL. PRL is a major factor in the proliferation and differentiation of breast epithelium and is essential for stimulation and maintenance of lactation. It has been also implicated in the development of breast cancer, tumor growth, and chemo-resistance. In the human, PRLR expression is controlled at the transcriptional level by multiple promoters (one generic, [PIII], and five human-specific [hPN1-hPN5]), which were defined and characterized in our laboratory. Each promoter directs transcription/expression of a specific non-coding exon 1 (E1-3, hEN1-hEN5), a common non-coding exon 2, and coding exons (E3-E11). The transcription of PRLR in breast cancer cells by the preferentially utilized PIII, which lacks an estrogen-responsive element, is enhanced by estradiol (E2)/ERa (estrogen receptor a) through complex formation with SP1 and C/EBPb, which associate with cognate elements at their DNA sites, inducing TFIIB and Pol II recruitment. Our studies revealed ERa constitutive homodimers (5). Complex formation of the ERa dimer (non DNA bound) with Sp1 and C/EBPbeta bound to their cognate sites is required for basal (constitutive ERa homodimers) and E2–induced transcriptional activation/expression of the human PRLR gene. PRL in tumoral breast causes cell proliferation by activating its cognate receptor. Exacerbation of PRL's actions in breast cancer resulting from increase receptor expression can explain resistance to estrogen inhibitors in breast cancer. Our studies reveal stimulation of PRLR transcription, mRNA, and protein in MCF7 cells by PRL in the absence of E2 abolished by mutation of a GAS site, Stat5 siRNA, or ER antagonist, indicating the participation of the ER in PRLR transcription via PRL/PRLR/Stat5. Increased recruitment of ER to Sp1 and C/EBP bound at their promoter sites is essential for PRL–induced receptor transcription. Our studies provide evidence for direct local actions of PRL independent of E2 in the up-regulation of PRLR transcription/expression via a Stat5/ER activation-loop. The studies are of relevance in states refractory to aromatase inhibitors, in which breast cancer progression could be fueled by endogenous prolactin.
Publications
- Liao M, Zhang Y, Kang JH, Dufau ML. Coactivator function of positive cofactor 4 (PC4) in Sp1-directed luteinizing hormone receptor (LHR) gene transcription. J Biol Chem 2011;286:7681-7691.
- Tsai-Morris CH, Sato H, Gutti R, Dufau ML. Role of gonadotropin-regulated testicular RNA helicase (GRTH/Ddx25) on polysomal associated mRNAs in mouse testis. PLoS One 2012;e32470:1-16.
- Villar J, Tsai-Morris CH, Dai L, Dufau ML. Androgen-induced activation of gonadotropin-regulated testicular RNA helicase (GRTH/Ddx25) transcription: essential role of a nonclassical androgen response element half-site. Mol Cell Biol 2012;32:1566-1580.
- Kavarthapu R, Tsai Morris CH, Fukushima M, Pickel J, Dufau ML. A 5' flanking region of gonadotropin-regulated testicular RNA helicase (GRTH/DDX25) gene directs its cell-specific androgen-regulated gene expression in testicular germ cells. Endocrinology 2013;154:2200-2207.
- Kang JH, Tsai-Morris CH, Dufau ML. Complex formation and interactions between transcription factors essential for human prolactin receptor gene transcription. Mol Cell Biol 2011;31:3208-3222.
Collaborators
- Sergio A. Hassan, PhD, Center for Molecular Modeling, Division of Computational Bioscience, NIH, Bethesda, MD
- James M. Pickel, PhD, Transgenic Core Facility, NIMH, Bethesda, MD
Contact
For more information, email dufau@helix.nih.gov.