

You are here: Home > Section on Formation of RNA
Physiological, Biochemical, and Molecular-Genetic Events Governing the Recognition and Resolution of RNA/DNA Hybrids

- Robert J. Crouch, PhD, Head, Section on Formation of RNA
- Susana M. Cerritelli, PhD, Staff Scientist
- Hyongi Chon, PhD, Postdoctoral Fellow
- Kiran Sakhuja, MS, MSc, Biologist
- Mariya London, BS, Technical Intramural Research Training Award
Damaged DNA is one of the leading causes of many human diseases and disorders. We study the formation and resolution of RNA/DNA hybrids, which occur during DNA and RNA synthesis. Such hybrid molecules may lead to increased DNA damage but may also play critical roles in normal cellular processes. We are interested in how RNA/DNA hybrids are resolved and in the role ribonucleases H (RNases H) play in their elimination. Two classes of RNases H are present in most organisms. Our studies showed that mice deleted for the Rnaseh1 gene arrest embryonic development at day 8.5 as a result of failure to amplify mitochondrial DNA. Others have found that the Aicardi-Goutières Syndrome (AGS), a severe neurological disorder with symptoms appearing at or soon after birth, can be caused by defective human RNase H2. We employ molecular-genetic and biochemical tools and yeast and mouse models in our research.
There are several types of RNA/DNA hybrids structures, which are formed and processed differently. Simple RNA/DNA hybrids consist of one strand of RNA paired with one strand of DNA. The HIV-AIDS reverse transcriptase (RT) forms such hybrids when copying its genomic RNA into DNA. The RT also has an RNase H domain that is structurally and functionally similar to the class I cellular RNase H and is necessary for several steps of viral DNA synthesis. R-loop hybrids have two separated DNA strands, with one hybridized to RNA while the other is in single-stranded form. The structures sometimes form during transcription and can lead to chromosomal breakage. However, they are also part of the normal recombination process of switching (recombination) from one form of immunoglobulin to another, resulting in different isoforms of antibodies. The first two types of hybrid are substrates for class I and II RNases H. The third is uniquely recognized by type 2 RNases H (Figure 1).
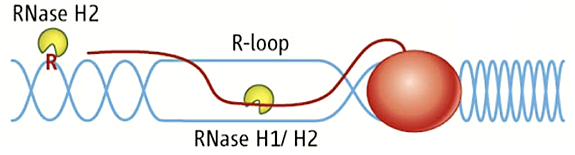
Figure 1. RNA/DNA hybrids and single rNMPs in DNA are substrates for RNase H2.
DNA is in blue, RNA is red line and letter R. The red oval represents RNA polymerase.
Contrasts between Class I and Class II RNases H
Previous Annual Reports detailed our work on RNase H1. RNase H1 recognizes the 2′-OH of four consecutive ribonucleotides while the DNA strand is distorted to fit in a pocket of the enzyme. Thus, the enzyme requires more than one ribonucleotide for cleavage of RNA in RNA/DNA hybrids. In both eukaryotes and prokaryotes, RNases H1 consist of a single polypeptide. In contrast, RNase H2 is a complex of three different polypeptides in eukaryotes but is a single polypeptide in prokaryotes. The catalytic subunit of the heterotrimeric RNase H2 of eukaryotes is similar in its primary amino acid sequence to the prokaryotic enzyme. RNase H2 can recognize and cleave a single ribonucleotide or the transition from the ribonucleotide in the case of RNA–primed DNA synthesis (e.g., rrrrrDDDD in DNA—italics indicate transition from ribonucleotide to deoxyribonucleotide—see Figure 1).
Dual activities of RNase H2 and Aicardi-Goutières syndrome
Eukaryotic RNases H2 recognize and resolve RNA hybridized or covalently attached to DNA—two chemically distinct structures (Figure 1; Cerritelli and Crouch, FEBS J 2009;276:1494; Reference 1)—using the same catalytic mechanism of hydrolysis. RNase H2 mutations that reduce catalytic activity or fail to properly interact with in vivo substrates cause AGS. Of the 29 different RNase H2 mutations associated with AGS, eight are in the A subunit, 14 in the B subunit, and 7 in the small C subunit (2). We previously expressed in E. coli and purified human RNases H2 with mutations corresponding to several seen in AGS patients. We found only one with significant loss of RNase H2 activity (3). Using the structure we determined, we located all known mutations in RNase H2 causing AGS (2). The wide distribution of the mutations suggests that modest changes in stability, interaction with other unknown proteins, and loss of catalysis can all cause AGS.
To distinguish the defects that persistent RNA/DNA hybrids and single ribonucleotides joined to DNA cause in vivo, we modified RNase H2 to make an enzyme that could only cleave one type of substrate. Based on a rational design comparing the structures of RNase H2 and RNase H3, we unlinked the two activities to yield an enzyme that processes RNA/DNA, but leaves single ribonucleoside monophosphates (rNMPs) attached to DNA. RNase H2 and RNase H3 have similar 3-D structures, but RNase H3 does not cleave single rNMPs in DNA. We first examined in vitro activities of our new RNase H2 mutant using, among others, the ribonucleotide excision repair (RER) assay recently developed by our collaborators Peter Burgers and Justin Sparks (4). The in vitro results show complete lack of removal of single ribonucleotides but only modest reduction in hydrolysis of RNA/DNA hybrids. In vivo, our RNase H2 mutant gave the signature 2–5 bp deletions in the yeast CAN1 gene associated with incorporation of ribos into DNA in the absence of RNase H2 (5). The mutant enzyme resolves RNA/DNA hybrids, which are substrates of both RNase H1 and RNase H2. However, our mutant RNase H2 identified a unique set of hybrids that formed when homologous recombination is defective owing to loss of SGS1 helicase, and that can only be processed by RNase H2, most likely because it has special access via contacts with other cell components. Thus, the synthetic defect observed in sgs1delta, rnh201delta strains results from problems associated with persistent R-loops. Given that our RNase H2 mutant enzyme resolves these R-loops, the synthetic defect is absent when the mutant RNase H2 is present in a strain deleted for SGS1.
The RNaseH2A G37S mutation, which is found in a few AGS patients, results in limited activity in vitro (3). We investigated how the corresponding mutation in yeast RNase H2 (G42S) protein functions. In vitro it had very poor activity on RNA/DNA hybrids and on cleavage of single rNMPs in DNA. In vivo it was almost as defective as a deletion of an RNase H2 subunit for removing single rNMPs (measured by 2–5 bp deletions in the CAN1 gene), had limited activity in removing R-loops accessible to either RNase H2 or RNase H1, yet displayed remarkably good activity when it interacted with other proteins involved in R-loops associated with DNA replication/repair.
We are developing mouse models of AGS to clarify which defects are associated with each RNase H2 activity. Mice bearing the RNase H2A G37S mutation in homozygous form are either stillborn or die within a few hours of birth. Mutations in another gene, TREX1, also cause AGS, and it has been shown that homozygous knockout (KO) mice are viable but die after a few weeks as a result of cardiomyopathy, which can be prevented by either blocking innate or adaptive immune responses. In contrast, Rnaseh2–KO mice die during embryogenesis, and the stillborn phenotype of the G37S mutation suggests a more severe defect than that seen in Trex1–KO mice. We attempted to rescue the stillborn phenotype by eliminating the innate or adaptive immune responses. G37S mice defective in Rag2 or INFRA1 are no different from the innate- or adaptive-competent mice, suggesting a different issue between the Trex1– and Rnaseh2–mutant mice.
Mitochondrial DNA and RNase H1 in mouse
During embryonic development, RNase H1 is required for progression beyond day E8.5. A single transcript of the mouse Rnaseh1 gene is translated to make two nearly identical proteins, one localized to the nucleus and the other to the mitochondrion. We previously showed that the Rnaseh1–deleted embryos fail to amplify mitochondrial DNA (mtDNA), causing developmental arrest. Nuclear DNA replicated normally in Rnaseh1–KO embryos. We are examining loss of the Rnaseh1 gene during B cell development to follow the process of RNase H1 depletion in a simpler system. Following conditional deletion of Rnaseh1 in mouse B cells at an early stage of B cell development, we find that resting, naive B cells are formed but are unable to become activated to class-switch to other isotypes (e.g, IgG), and sera from the mice have a major deficit in antibodies. We are currently determining whether the loss of mtDNA is the explanation for the inability of the resting B cells to be completely activated. The fact that loss of the “housekeeping” Rnaseh1 gene permits extensive development is in sharp contrast to the finding that deletion of the La SSB gene (another “housekeeping gene”), at the same stage of B cell development, results in complete absence of any B cells at the stage at which the gene is deleted (Gaidamakov et al., Mol Cell Biol 2013;in press) (see report from Richard Maraia).
In collaboration with Ian Holt, we are examining the roles of RNase H1 in mitochondrial DNA replication by, among other techniques, analyzing intermediates on two-dimensional gels. Our findings thus far indicate that elevated expression of RNase H1 in mitochondria alters mtDNA replication. In addition, data obtained in Holt's laboratory by John Holmes supporting the existence of RNA/DNA hybrids as replication intermediates indicate a bi-directional mode of DNA replication for this organelle.
Additional Funding
- Scientific Directors Intramural Award
Publications
- Cerritelli SM, Chon H, Crouch, RJ. A new twist for topoisomerase. Science 2011;332:1510-1511.
- Figiel M, Chon H, Cerritelli SM, Cybulska M, Crouch RJ, Nowotny M. The structural and biochemical characterization of human RNase H2 complex reveals the molecular basis for substrate recognition and Aicardi-Goutières Syndrome defects. J Biol Chem 2011;286:10540-10550.
- Chon H, Vassilev A, DePamphilis ML, Zhao Y, Zhang J, Burgers PM, Crouch RJ, Cerritelli SM. Contributions of the two accesory subunits, RNASEH2B and RNASEH2C, to the activity and properties of the human RNase H2 complex. Nucleic Acids Res 2009;37:96-110.
- Sparks JL, Chon H, Cerritelli SM, Kunkel TA, Johansson E, Crouch RJ, Burgers PM. RNase H2-initiated ribonucleotide excision repair. Mol Cell 2012;47:980-986.
- Chon H, Spark JL, Rychlik M, Nowotny M, Burgers PM, Crouc RJ, Cerritelli SM. RNase H2 roles in genome integrity revealed by unlinking its activities. Nucleic Acids Res 2013;41:3130-3143.
Collaborators
- Peter Burgers, PhD, Washington University, St. Louis, MO
- Ian Holt, PhD, MRC-Dunn Nutrition Unit, Cambridge, UK
- Richard J. Maraia, MD, Program in Genomics of Differentation, NICHD, Bethesda, MD
- Herbert C. Morse, MD, Laboratory of Immunopathology, NIAID, Bethesda, MD
- Roger Woodgate, PhD, Laboratory of Genomic Integrity, NICHD, Bethesda, MD
Contact
For more information, email crouch@helix.nih.gov or visit sfr.nichd.nih.gov.