

You are here: Home > Section on Molecular Genetics of Immunity
Gene Regulation in Innate Immunity

- Keiko Ozato, PhD, Head, Section on Molecular Genetics of Immunity
- Anup Dey, PhD, Biologist
- Tomohiko Kanno, MD, PhD, Staff Scientist
- Natarajan Ayithan, PhD, Visiting Fellow
- Monica Gupta, PhD, Visiting Fellow
- Rui Kamada, PhD, Visiting Fellow
- Naoyuki Sarai, PhD, Visiting Fellow
- Yuko Yoshida, PhD, Visiting Fellow
- Hiroaki Yoshii, PhD, Visiting Fellow
- Tamaki Ito, PhD, JSPS-NIH Fellow
- Ankur Narain, BS, Postbaccalaureate Fellow
Macrophages (MФ) and dendritic cells (DC) respond to pathogen stimuli by producing cytokines, including interferons (IFNs), IL-1, IL-6, and TNF-alpha, to impart anti-viral and anti-microbial status to the host. Our goal is to study the molecular pathways that direct the development and function of MФ and DC. Our long-term interest has been the role of the transcription factor IRF8 in innate immunity. IRF8 is expressed in MФ and DC at high levels and is required for the production of both type I and type II IFNs. IRF8 is essential for mounting the first line of defense against various invading pathogens prior to the initiation of antigen-specific immune responses. Autophagy is one of mechanisms by which MФ and DC eliminate pathogens. It is thought that autophagy plays a role in host resistance against HIV-1 infection. We are interested in the role of IRF8 in autophagy and innate immune responses.
Transcriptionally active genes are embedded in chromatin that is dynamically exchanged, whereas silenced genes are surrounded by more stable chromatin. The chromatin environment influences transcriptional processes and controls epigenetic regulation. We are interested in BRD4, a chromatin-binding protein associated with transcribed genes. We are also interested in histone H3.3, the variant histone that is selectively associated with actively expressed genes. BRD4 is a 200 kDa nuclear protein carrying two tandem bromodomains through which it binds to acetylated chromatin. BRD4 also interacts with the elongation factor P-TEFb and regulates transcription of many genes, including those induced by external stimuli. BRD4 is implicated in transcriptional memory across cell division because it stays on condensed chromosomes during mitosis and affects gene expression in daughter cells. Despite close structural similarity with the standard H3.1 and H3.2 histones, histone H3.3 has an extraordinary property in that it is incorporated into nucleosomes and DNA only in actively transcribed genes. In contrast, H3.1, H3.2, and other standard core histones are incorporated into nucleosomes during DNA replication. The difference between H3.3 and H3.1/2 reflects distinct chromatin activities during replication and transcription. Although there is mounting recognition of the importance of transcription-coupled histone incorporation, the process and its physiological significance are still shrouded in mystery. Our goal is to elucidate the activity of BRD4 and histone H3.3 in the context of transcriptional activation and epigenetic memory.
IRF8 is a master regulator of autophagy genes required for autophagosome-lysosomal functions.
Autophagy is a conserved catabolic process active in most cells and is designed to eliminate misfolded and damaged self proteins. The process is essential during embryonic development: mice with the disrupted autophagy gene Atg5 are embryonic lethal. MФ and DC employ autophagy as a mechanism of host resistance against invading pathogens. Autophagy is activated by pathogen infection and by IFNγ in MФ, which trigger a cascade of events leading to autophagosme formation, fusion with lysosomes, and protein degradation via ubiquitin conjugation and sequestration with SQSTM1 (p62). The processes are regulated by more than 20 genes that are specialized in executing autophagy. While autophagy helps to clear invading pathogens, some pathogens utilize autophagy to enhance their growth and survival. Through microarray and ChIP-on-chip analyses on IRF8+/+ and IRF8−/− DC, we found that IRF8 regulates expression of some autophagy-related genes. The extension of these analyses led to the observation that IRF8 stimulates at least 17 autophagy genes in MФ. Further tests found that of the 17 genes, 12 could be rescued for expression in IRF8−/− MФ upon IRF8 transfer. For seven of these genes, IRF8 bound to the promoter regions, demonstrating that IRF8 is a major regulator of autophagy gene expression. In accordance, IRF8−/− MФ are defective in autophagosome formation, the process critical for capturing of misfolded proteins. We found IRF8−/− MФ to be also defective in lysosomal fusion required for final degradation of the captured proteins. To summarize, IRF8 is required for full autophagy function in MФ and DC, thereby critically contributing to innate immunity.
Histone methyltransferase WHSC1 directs H3.3 incorporation in activated genes by recruiting the chromatin assembly factor HIRA.
H3.3 is very similar to the canonical histone H3 (H3.1 and H3.2) in its 3-D structure, differing only in a few amino acids. Unlike the canonical histone H3, however, H3.3 (encoded by two genes) is synthesized outside the S phase and is incorporated into chromatin during transcription. It is thought that H3.3 associates with the preexisting canonical histone H4 to form a new nucleosome, presumably along with H2AZ and the preexisting H2B. H3.3 incorporation into chromatin requires specific chromatin-assembly factors distinct from those involved in replication-coupled histone deposition. HIRA is a major H3.3–specific histone chaperone dedicated to transcription-coupled H3.3 incorporation. HIRA carries the WD40 domain and B domain, through which it interacts with three other subunits to act as a chromatin-assembly factor. Another H3.3–specific chaperone, ATRX, is reported to be involved in H3.3 incorporation into the telomeres.
We are interested in transcription-coupled H3.3 incorporation. During the past year, we studied the mode and kinetics of H3.3 deposition during and after transcriptional activation. Analyzing the interferon (IFN)–stimulated genes (ISGs) Ifit1, Mx1, Oas1, and Stat1 as a model, we showed that H3.3 is incorporated into all four ISGs as soon as their transcription begins. Notable features of IFN–induced H3.3 deposition were that H3.3 deposition displayed a sharp positional gradient, in that the deposition was the greatest in the 3′ transcription end site, and that H3.3 deposition continued long after transcription had ceased, even 48 h after IFN stimulation. Essentially, an identical pattern of H3.3 deposition was seen for the c- Fos and c- Jun genes, which encode proteins that form the heterodimeric transcription factor AP-1 after UV stimulation. We noted that trimethylation of H3K36 was induced after stimulation and that the temporal and positional patterns of increase closely correlated with H3.3 incorporation. Methylation of H3K36 is catalyzed by the SET2 family of histone methylases, including WHSC1 (also known as NSD2). We investigated whether WHSC1 plays a role in transcription-coupled H3.3 deposition by testing Whsc1−/− mouse embryonic fibroblasts (MEFs). We found that IFN–induced H3.3 deposition was completely absent in Whsc1−/− MEFs, while Whsc1+/+ MEFs supported the expected H3.3 deposition. We found that WHSC1 interacts with the chromatin-binding factor BRD4 and is thus critical for the recruitment of the transcription elongation factor P-TEFb. In the absence of WHSC1, ISG elongation was strongly impaired, resulting in severe reduction of ISG mRNA induction. Supporting the role of WHSC1 in elongation, the elongating form of RNA polymerase II (S2Pol II) was present on the ISGs at very low levels in Whsc1−/− cells. Chromatin immunoprecipitation (ChIP) analysis demonstrated that WHSC1 was recruited to the ISGs upon IFN stimulation. Distributed throughout the coding regions, WHSC1 remained on the ISGs long after the cessation of ISG transcription. In parallel with WHSC1 recruitment, HIRA bound to the ISGs after IFN stimulation and remained on ISGs as long as WHSC1 did. Furthermore, WHSC1 directly interacted with HIRA through the internal PHD domain and the HMG DNA–binding domain, independently of the interaction with BRD4. Consistent with the interaction of WHSC1 and HIRA, WHSC1 and HIRA bound to the ISGs as a protein complex, as confirmed by re-ChIP assays. Taken together, our studies reveal two phases of H3.3 deposition: an initial elongation-coupled phase of deposition, which is then switched to the post-transcription phase of deposition. It is clear that WHSC1 plays a central role in both phases.
Construction of H3.3–HA knock-in mice and validation of the experimental model
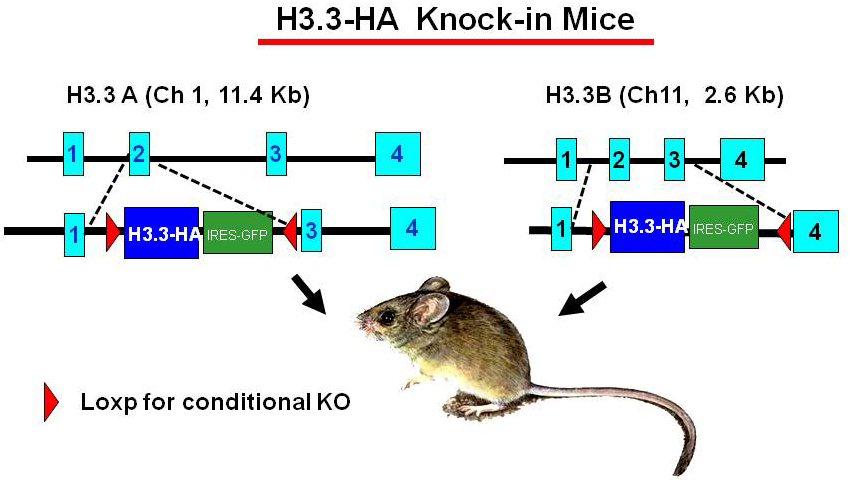
Click image to enlarge.
Figure 1. H3.3–HA knock-in mice
To study the activity of H3.3 in vivo, we generated mouse strains in which the endogenous H3.3 genes (H3.3A, H3f3a and H3.3B, H3f3b) are replaced by hemagglutinin (HA)-tagged H3.3 genes (Figure 1). The knock-in targeting vectors contain HA–tagged H3.3 cDNA (blue) and an IRES–driven GFP (green) to allow independent monitoring of the expression of the inserted genes. The inserted H3.3A–HA and H3.3B–HA fragments are flanked by the LoxP sites (red) to allow conditional knockout of the genes. The original targeting vectors had the neomycin- and hygromycin-resistant genes for ES cell selection but were removed by FRT–based recombination to avoid potential adverse effects in the mouse. Heterozygous mice for the H3.3A–HA and H3.3B–HA loci are viable and fertile, although the birth rate of homozygous mice appears significantly lower than that expected for Mendelian inheritance. We confirmed that H3.3–HA is expressed at variable levels throughout adult tissues. Preliminary ChIP analysis performed with anti–HA antibody indicated that H3.3–HA is deposited in induced genes in bone marrow–derived macrophages stimulated with IFNg and toll-like receptor (TLR) signaling. We established mouse embryonic fibroblasts (MEFs) that carry homozygous and heterozygous H3.3A–HA and H3.3B–HA and we plan to analyze the MФ and MEFs for global H3.3 deposition, using the ChIP-seq approach.
The role of IRF8 in protection against infectious pathogens
IRF8, a transcription factor of the IRF family, is highly expressed in MФ and DC and is shown to be important for host resistance against various pathogens including Mycobacterium tuberculosis, Salmonella, and Listeria, as well as against RNA and DNA viruses. We have been interested in Ebola viruses (EBOV), which cause severe hemorrhagic fever with high fatality, given that the viruses infect MФ and DC as the initial site of infection and inhibit IFN induction and IFN signaling. EBOV virus-like particles (VLPs), genome-free viral proteins, composed of the matrix protein (VP40), membrane glycoprotein, and nucleoprotein (NP), serve as a promising vaccine candidate against EBOV infection. When given prior to EBOV infection, VLPs fully protect rodents and nonhuman primates from the EBOV disease. Recent studies show that, even when given following lethal EBOV infection, EBOV can protect mice. We investigated the mechanisms by which VLPs provide post-exposure protection, focusing on the role of IFN and IRF8. We found that VLP administration fails to protect Ifnar−/− mice, which lack type I interferon (IFN) signaling. In wild-type mice, VLPs accelerated induction of many IFN–stimulated genes (ISGs) in liver and spleen. The ISGs included classical anti-viral proteins and negative feedback factors known to restrict excessive inflammatory responses. However, VLPs did not induce ISGs in Ifnar−/− mice, and consequently uncontrolled viral growth and elevated proinflammatory cytokines and chemokines ensued in the mice, indicating that type I IFNs broadly attenuate inflammatory responses through IFN–induced negative feedback factors. We also found that VLPs were unable to establish preventive anti–EBOV immunity in Irf8−/− mice, illustrating the role of IRF8 in initial IFN activation in critical cell types. Supporting the in vivo results, we showed that VLPs activate type I IFN genes and ISGs through the toll-like receptor signaling in MФ and DC. Together, VLPs afford post-exposure protection by expeditious induction of type I IFNs in the host.
Additional Funding
- Trans NIH-FDA Biodefense Program; IATAP Program
Publications
- Nishiyama A, Dey A, Tamura A, Ko, M, Ozato K. Activation of JNK triggers release of Brd4 from mitotic chromosomes and mediates protection from drug-induced mitotic stress. PLoS One 2012;7(5):e34719.
- Chang T-H, Xu S, Tailor P, Kanno T, Ozato K. The SUMO deconjugating enzyme SENP1 switches IRF8 from a repressor to an activator and promotes macrophage activation. J Immunol 2012;189:3548-3556.
- Patel MC, Debrosse M, Smith M, Dey A, Huynh W, Heightman TD, Tamura T, Ozato, K. BRD4 recruits the pausing complex NELF/DSIF along with P-TEFb to coordinate transcription elongation from interferon stimulated genes. Mol Cell Biol 2013;33:2497-2507.
- Ayithan N, Bradfute SB, Anthony SM, Stuthman KS, Dye JM, Bavari S, Bray M, Ozato. K. Ebola virus-like particles stimulate type I interferons and proinflammatory cytokine expression through the toll-like receptor and interferon signaling pathways. J Interferon Cytokine Res 2013;Epub ahead of print.
- Sarai N, Nimura. K, Tamura T, Kanno T, Patel MC, Heightman TD, Ura K, Ozato, K. WHSC1 links transcription elongation to HIRA mediated histone H3.3 deposition. EMBO J 2013;32:2392-2406.
Collaborators
- Steven B. Bradfute, PhD, United States Army Medical Institute of Infectious Diseases, Fort Detrick, MD
- Ben-Zion Levi, PhD, Technion, Israel Institute of Technology, Haifa, Israel
- Herbert Morse II, MD, Laboratory of Immunopathology, NIAID, Rockville, MD
- Tomohiko Tamura, MD, PhD, Tokyo University, Tokyo, Japan
Contact
For more information, email ozatok@mail.nih.gov or visit ozatolab.nichd.nih.gov.