Regulation of Childhood Growth

- Jeffrey Baron, MD, Head, Section on Growth and Development
- Kevin Barnes, PhD, Senior Research Assistant
- Julian Lui, PhD, Staff Scientist
- Youn Hee Jee, MD, Staff Clinician
- Benjamin Hauser, BS, Postbaccalaureate Fellow
- Adrian Temnycky, BS, Postbaccalaureate Fellow
- Elaine Zhou, BA, Postbaccalaureate Fellow
Children grow taller because their bones grow longer. Bone elongation occurs at the growth plate, a thin layer of cartilage found near the ends of juvenile bones. In the growth plates, new cartilage is produced through chondrocyte proliferation, hypertrophy, and cartilage matrix synthesis, and then the newly formed cartilage is remodeled into bone. The process, termed endochondral ossification, results in bone elongation, which causes children to grow in height (linear growth). Consequently, mutations in genes that regulate growth-plate chondrogenesis cause abnormal bone growth and short stature in children. Depending on the severity and nature of the genetic abnormality, the phenotype can range from chondrodysplasias with short, malformed bones, to severe, often disproportionate, short stature, to mild proportionate short stature. If the genetic defect affects tissues other than the growth plate cartilage, the child may present with a more complex syndrome that includes other clinical abnormalities.
We investigate the cellular and molecular mechanisms governing childhood growth and development. We focus on growth at the growth plate, which drives bone elongation and therefore determines height. One goal of this work is to gain insight into the many human genetic disorders that cause childhood growth failure or overgrowth. A second goal is to develop new treatments for children with severe growth disorders.
Novel genetic causes of childhood growth disorders
For many children who are brought to medical attention for linear growth disorders, clinical, laboratory, and genetic evaluation fails to identify the underlying etiology. Genome-wide association studies and molecular studies on growth-plate biology suggest that there are hundreds of genes that control linear growth. Therefore, it is likely that many genetic causes of linear growth disorders remain to be discovered.
To discover new genetic causes of childhood growth disorders, we invite families with monogenic growth disorders to the NIH Clinical Center, where we evaluate the clinical, biochemical, and radiological features of the condition. We then obtain DNA samples from informative family members and use powerful genetic approaches, including SNP arrays, to detect deletions, duplications, mosaicism, and uniparental disomy, combined with exome sequencing to detect single-nucleotide variants and small insertions/deletions in coding regions and splice sites. When sequence variants that are likely to cause the disorder are identified, we study the variants and the genes in which they occur in the laboratory to confirm that the variant is pathogenic, to elucidate the pathogenesis of the disorder, and to explore the role of the gene in normal growth.
We have used this approach to identify new causes of childhood growth disorders. For example, we previously found that variants in aggrecan (ACAN), a component of cartilage extracellular matrix, cause autosomal-dominant short stature with advanced skeletal maturation and that such patients also tend to develop early-onset osteoarthritis. We also found evidence that heterozygous deletion of CYP26A1 and CYP26C1, which encode enzymes that metabolize retinoic acid (RA), cause elevated RA concentrations, which accelerate bone and dental maturation in humans and cause developmental defects involving the eye and central nervous system. In additional studies, we found that variants in QRICH1, a gene of unknown function, cause a chondrodysplasia resulting from impaired growth-plate chondrocyte hypertrophic differentiation, in addition to the previously described developmental delay [Reference 1].
Figure 1. Pedigree of a family with markedly delayed puberty
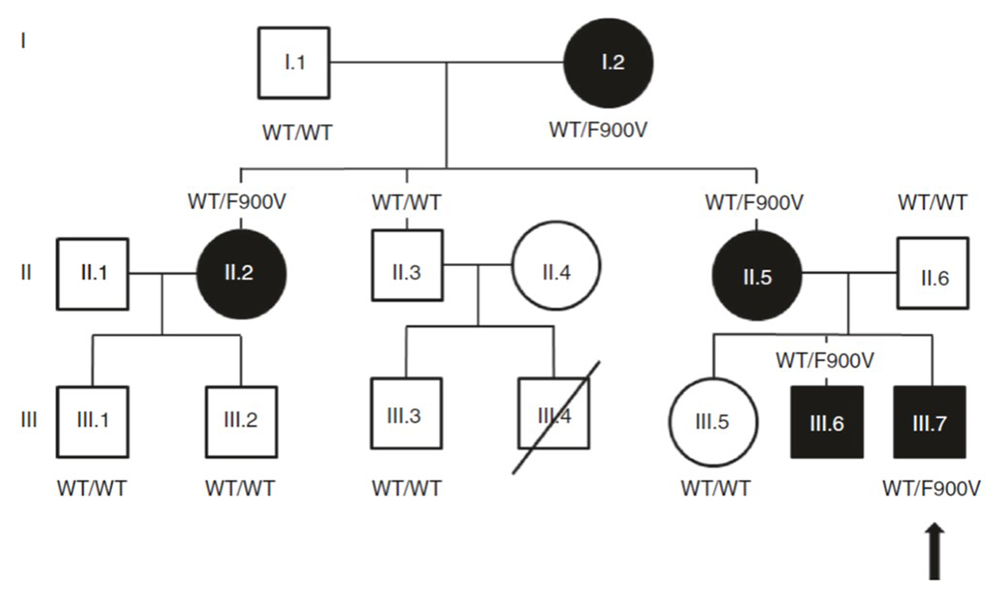
Click image to view.
Arrow indicates proband. Closed symbol, delayed puberty; open symbol, normal timing of puberty; F900V, a variant in DLG2/PSD-93; WT, wild-type DLG2/PSD-93.
Recently, we applied this approach to evaluate a child with markedly delayed puberty, which was inherited as an autosomal dominant trait (Figure 1). Exome sequencing identified a rare missense variant (F900V) in DLG2, which co-segregated precisely with the delayed puberty phenotype in the extended family. DLG2 encodes PSD-93, an anchoring protein of N-methyl-D-aspartate (NMDA) receptors. Interestingly, NMDA receptors have been shown to control the timing of puberty in laboratory animals. We studied the impact of the identified variant in a GnRH neuronal cell line. GnRH neurons play a central role in the initiation of puberty. We found evidence that the variant impairs binding of PSD-93 to FYN (a protein tyrosine kinase), which diminishes phosphorylation and activation of NMDA receptors, leading to reduced GnRH expression (Figure 2). Next, we identified sequence variants in DLG2/PSD-93 in three subjects with isolated hypogonadotropic hypogonadism, a condition that leads to a failure of pubertal development. The variants also diminished binding to FYN and reduced Gnrh1 expression in vitro. The findings provide evidence that variants in DLG2 cause delayed puberty and contribute to isolated hypogonadotropic hypogonadism by a mechanism that involves impaired binding to Fyn, diminished NMDA receptor phosphorylation and signaling, and consequently decreased GnRH expression [Reference 2].
Figure 2. Effect of F900V DLG2/PSD-93 variant on Gnrh1 expression in GT1-7 cells
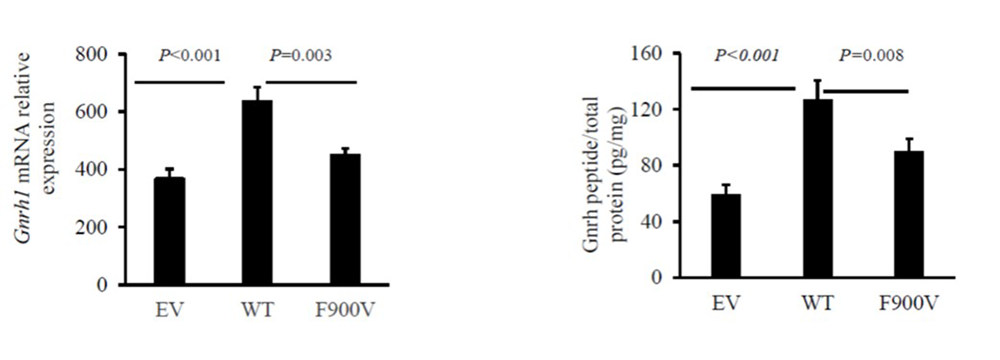
Click image to view.
In GT1-7 cells (a mouse hypothalamic GnRH neuronal cell line), wild-type (WT) PSD-93 expression stimulated Gnrh1 expression compared with empty vector (EV) for both mRNA (left panel) and GnRH peptide expression (right panel). The F900V PSD-93 mutant impaired the stimulation both in mRNA levels (left panel) and intracellular peptide levels (right panel).
Molecular and cellular mechanisms by which specific genes and pathways regulate childhood growth
Our group also studies the fundamental mechanisms governing skeletal growth. For example, we previously studied the role of EZH2 in the growth plate. EZH2 encodes a histone methyltransferase that catalyzes the trimethylation of histone H3 at lysine 27 (H3K27), which serves as an epigenetic signal for chromatin condensation and transcriptional repression. We found that loss of EZH1 and EZH2 in mice impaired bone growth, and we explored the cellular and molecular mechanisms involved. We also previously investigated the mechanisms that cause bones at different anatomical locations to vary dramatically in size and found that the postnatal developmental program that normally causes bone growth to decelerate with age plays out more rapidly in small bones than in large bones.
Recently, we studied the role of midkine in human growth, development, and disease. Midkine is a heparin-binding growth factor, which is highly expressed in several organs during embryogenesis. We previously found that it is present in high concentrations in human amniotic fluid and also in biopsy specimens of specific malignancies. We next analyzed plasma midkine concentrations in healthy, normal-weight children, healthy adults, obese children, girls and young women with anorexia nervosa, and children with idiopathic short stature. In healthy children, we found that plasma midkine concentrations declined with age. The decline occurred primarily during the first year of life. Plasma midkine did not significantly differ between males and females or between race/ethnic groups. Midkine concentrations were not correlated with body mass index (BMI) standard deviation score (SDS), fat mass (kg), or percent total body fat assessed by DXA (dual-energy X-ray absorptiometry); we found no difference in midkine between children with anorexia nervosa, healthy weight, and obesity. For children with idiopathic short stature, midkine concentrations did not differ significantly from normal-height subjects. The findings provide useful reference data for studies of plasma midkine in children with malignancies and other pathological conditions [Reference 3].
New treatment approaches for growth plate disorders
Recombinant human growth hormone (GH) is commonly used to treat short stature in children. However, GH treatment has limited efficacy, particularly in severe, non-GH–deficient conditions such as chondrodysplasias, and has potential off-target effects. Systemic insulin-like growth factor 1 (IGF-1) treatment has similar deficiencies. There are several endocrine and paracrine factors that promote chondrogenesis at the growth plate, which could potentially be used to treat such disorders. Targeting the growth factors specifically to the growth plate might augment the therapeutic skeletal effect, while diminishing undesirable effects on non-target tissues. To develop growth plate–targeted therapy, we previously used yeast display to identify single-chain human antibody fragments that bind to cartilage with high affinity and specificity. As a first test of the approach, we created fusion proteins combining the cartilage-targeting antibody fragments with IGF-1, an endocrine/paracrine factor that positively regulates chondrogenesis. Such fusion proteins retained both cartilage binding and IGF-1 biological activity, and they were able to stimulate bone growth in an organ culture system. Using a GH–deficient mouse model, we found that subcutaneous injections of the fusion proteins increased growth plate height without increasing proliferation in kidney cortical cells, demonstrating greater on-target efficacy at the growth plate and less off-target effect on the kidney than does IGF-1 alone. Our findings provide proof of principle that targeting therapeutics to growth plate cartilage could improve treatment for childhood growth disorders [Reference 4].
We are currently applying the approach to target other endocrine and paracrine factors that stimulate chondrogenesis to the growth plate. We are exploring the utility of the approach to both stimulate growth-plate chondrogenesis non-specifically and also reverse specific genetic defects in growth-plate function by modulating the abnormal molecular pathway responsible for the growth failure.
Additional Funding
- NIH U01 award: 1U01HD086838-01A1 (2017-2021, ongoing): “Genetic Diagnosis of Childhood Growth Disorders”
Publications
- Lui JC, Jee YH, Lee A, Yue S, Wagner J, Donnelly D, Vogt K, Baron J. QRICH1 mutations cause a chondrodysplasia with developmental delay. Clin Genet 2019;95:160-164.
- Jee YH, Won S, Lui JC, Jennings M, Whalen P, Yue S, Temnycky AG, Barnes KM, Cheetham T, Boden MG, Radovick S, Quinton R, Leschek EW, Aguilera G, Yanovski JA, Seminara SB, Crowley WF, Delaney A, Roche KW, Baron J. DLG2 variants in patients with pubertal disorders. Genet Med 2020;22:1329-1337.
- Jee YH, Lee KS, Yue S, Leschek EW, Boden MG, Jadra A, Klibanski A, Vaidyanathan P, Misra M, Chang YP, Yanovski JA, Baron J. Plasma midkine concentrations in healthy children, children with increased and decreased adiposity, and children with short stature. PLoS One 2019;14:e0224103.
- Lui JC, Colbert M, Cheung CS, Ad M, Lee A, Zhu Z, Barnes K, Dimitrov DS, Baron J. Cartilage-targeted IGF-1 treatment to promote longitudinal bone growth. Mol Ther 2019;27:673-680.
- Lui JC, Yue S, Lee A, Kikani B, Temnycky A, Barnes KM, Baron J. Persistent Sox9 expression in hypertrophic chondrocytes suppresses transdifferentiation into osteoblasts. Bone 2019;125:169-177.
Collaborators
- Greti Aguilera, MD, Scientist Emeritus, NICHD, Bethesda, MD
- Angela Delaney Freedman, MD, St. Jude Children’s Research Hospital, Memphis, TN
- Lijin Dong, PhD, Genetic Engineering Core, NEI, Bethesda, MD
- Ellen Leschek, MD, Division of Diabetes, Endocrinology, and Metabolic Diseases, NIDDK, Bethesda, MD
- Thomas Markello, MD, PhD, Undiagnosed Diseases Program, NHGRI, Bethesda, MD
- Madhusmita Misra, MD, Harvard Medical School, Massachusetts General Hospital, Boston, MA
- Ola Nilsson, MD, PhD, Karolinska Institute, Stockholm, Sweden
- Sally Radovick, MD, Rutgers Biomedical and Health Sciences, Robert Wood Johnson Medical School, New Brunswick, NJ
- Katherine W. Roche, PhD, Receptor Biology Section, NINDS, Bethesda, MD
- Jack Yanovski, MD, PhD, Section on Growth and Obesity, NICHD, Bethesda, MD
Contact
For more information, email jeffrey.baron@nih.gov or visit http://baron.nichd.nih.gov.