Receptors and Actions of Peptide Hormones and Regulatory Proteins in Endocrine Mechanisms

- Maria L. Dufau, MD, PhD, Head, Section on Molecular Endocrinology
- Raghuveer Kavarthapu, PhD, Staff Fellow
- Muruganath Kumar Raju, PhD, Postdoctoral Fellow
- Rajakumar Anbazhagan, PhD, Visiting Fellow
The Section investigates the molecular basis of peptide hormones and their receptors in the control of gonadal function. Studies encompass the structure and transcriptional regulation of the luteinizing-hormone (LH) and prolactin receptors. We study the regulatory mechanism(s) involved in the progression of spermatogenesis and the control of Leydig cell function. We investigate novel gonadotropin-regulated genes relevant to the progression of testicular gametogenesis, Leydig cell function, and other endocrine processes. Our studies concentrate on the function and regulation of the gonadotropin-regulated testicular RNA helicase (GRTH/DDX25), a member of the DEAD-box family of RNA helicases discovered, cloned and characterized in our laboratory, which is essential for the progress of spermatogenesis. The various functions of GRTH/DDX25 provide fertile ground for the development of a non-hormonal male contraceptive.
Gonadotropin-regulated testicular RNA helicase
GRTH/DDX25 is present in Leydig cells (LC), meiotic (pachytene spermatocytes), and haploid germ cells (round and elongated spermatids) [Reference 1]. Males lacking GRTH are sterile owing to azoospermia, which results from failure of round spermatids (RS) to elongate. We demonstrated its participation in the nuclear export/transport of specific mRNAs, the structural integrity of the chromatoid body (CB), and in storage/processing of relevant mRNAs and their transit to and association with actively translating polyribosomes, where GRTH may regulate translational events. GRTH is the only member of the DEAD-box family regulated by hormones. GRTH transcription is stimulated in LCs by LH/cAMP through direct actions of androgen (A) and the A receptor (AR) (autocrine), and indirectly in germ cells (paracrine) through the AR in Sertoli cells. The upstream region of the GRTH gene directs its expression in germ cells and downstream in the LC. Through these regions, A/AR exerts their direct (endogenous) regulation of the GRTH gene in LC and indirectly in germ cells. Functional binding sites for germ-cell nuclear factor (GCNF) present in RS and spermatocytes (SP) and its regulation by A/AR were identified in the distal region of the GRTH gene, operative selectively in RS. Current knowledge indicates actions of A on GCNF cell-specific regulation of GRTH expression in germ cells (RS). Also, we previously established that GRTH exerts negative autocrine regulation of GCNF, linking A actions to germ cells through GCNF as an A–regulated transfactor that controls transcription/expression of GRTH, providing a connection between androgen action and two relevant germ-cell genes (GRTH and GCNF), which are essential for the progress of spermatogenesis, and establishing their regulatory interrelationship.
Our early studies revealed that missense mutation of R to H at amino acid 242 of GRTH, found in 5.8% of patients with complete loss of sperm, causes loss of the 61 kDa phospho-species (pGRTH), with preservation of the 52 kDa nonphospho form. The finding provided an avenue through which to elucidate the function of pGRTH in spermatogenesis. We generated humanized mutant GRTH knock-in (KI) mice [Reference 2]. The mice are sterile, with reduced testicular size and lack of sperm (arrest at step 8 of RS), and demonstrate complete loss of the pGRTH species but with preservation of the nuclear 52 kDa form. The mouse model permits the study of the biological/biochemical functions of the cytoplasmic pGRTH. In KI mice, the nuclear export transport and functions of GRTH are preserved (i.e., mRNA export, miRNA regulation), while the cytoplasmic functions, including shuttling of messages, storage in the CB, and translational events all requiring pGRTH, are absent. We observed marked reduction in the size of the CB in RS and lack of pGRTH in the CB. Germ cell apoptosis was present in pachytene spermatocytes (PS) and RS. In contrast to knockout (KO) mice, KI mice showed no changes in miRNA biosynthesis, which excludes participation of pGRTH as transcriptional regulator of the microprocessor complex (the RNase III Drosha and the RNA–binding protein DCGR), which affects primiRNAs formation, but indicates the participation of non-phospho GRTH in these processes. In KI mice, there is loss of chromatin remodeling and of related proteins, including TP2, PRM2, and TSSK6. Significant decreases in their mRNA and half-lives indicate that their association with pGRTH in the cytoplasm protects such mRNAs from degradation. Also, our previous work showed that pGRTH stimulates TP2 translation in a 3′ UTR–dependent manner.
In recent studies, we elucidated the GRTH phospho-site at a threonine (Tr239), which is structurally adjacent to the mutant site found in patients (R242H) [Reference 3]. Molecular modelling of the phospho-site, based on the RecA domain 1 of the DDX19 (an RNA helicase) crystal structure, pointed to the amino acids that formed the GRTH/PKA interface, solvent accessibility, and H-bonding. In addition to the core residues T239 and R242, these include amino acids E165, K240, and D237. The relevance of these residues was demonstrated by single or double mutations that caused reduction in or abolition of the p-GRTH at Tr239. pGRTH is the cytoplasmic form, essential for the progress of spermatogenesis beyond step 8 of round spermatids, and consequent lack of sperm formation [Reference 4]. The deleterious effects on GRTH phosphorylation caused by the mutations did not result from changes of protein kinase A alpha (PKAa) catalytic binding affinity but rather from consequential structural changes that can affect PKA catalytic efficiency. Studies based on the abolition of the phospho-form provide the basis for drug design and virtual and throughput screening for discovery of a reversible chemical inhibitor for use as male contraceptive. During the past year, we determined that cyclic peptides that fit the shallow pocket of GRTH/protein kinase A are preferred compounds to block GRTH phosphorylation and amenable for use in the development of oral non-hormonal male contraceptives.
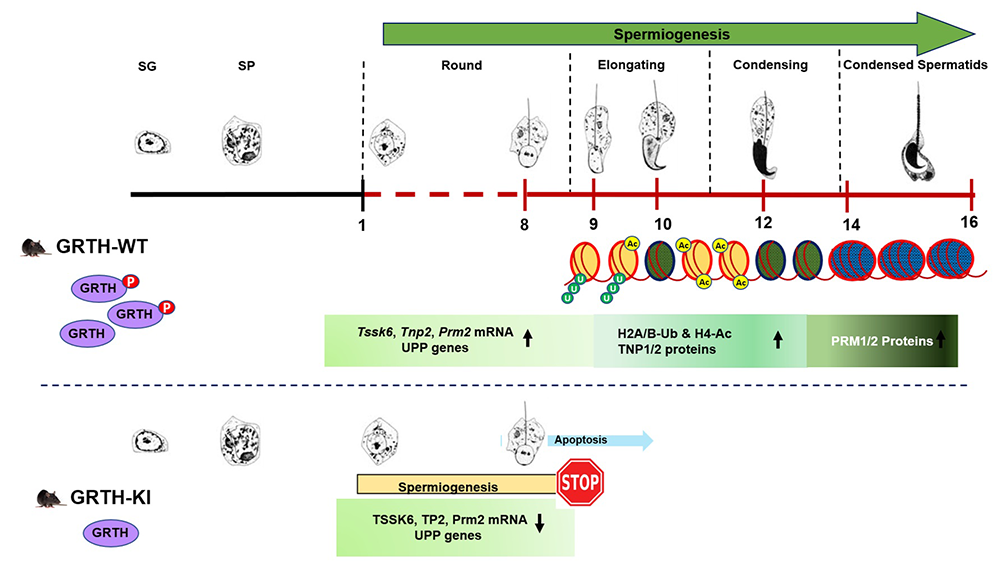
Work on transcriptome analysis of mice germ cells using RNA-Seq provided further insights regarding linking pGRTH to histone ubiquitination and acetylation, essential for chromatin compaction and spermatid development during spermiogenesis [Reference 4] (Figure 1). We also initiated studies on the role of phosphorylated GRTH in the storage of messages in the CB, an organelle present in germ cells and found to be a key site for the storage of mRNAs of relevance for the progress of spermatogenesis.
Figure 1. Role of p-GRTH in histone ubiquitination and acetylation essential for round spermatid development during spermiogenesis.
Click image to view.
Schematic diagram showing progression of mice spermiogenesis, during which germ cells undergo 16 different steps of development. In WT mice, during the process of spermiogenesis in round spermatids (RS), we observed increased and stable expression of Tssk6, Tnp2, Prm2, and UPP mRNAs until ready for translation in step 9 of RS. During spermatid elongation, histones undergo ubiquitination (H2A/B-Ub) and acetylation (H4-Ac), resulting in their removal and replacement with transition proteins and protamines, making the chromatin more compact/condensed. Of note is that, in KI mice, loss of p-GRTH has a direct impact on the expression of UPP genes required for ubiquitination and subsequent acetylation, which impairs round spermatids to elongation [Reference 4].
Publications
- Dufau ML, Kavarthapu R. Gonadotropin regulated testicular RNA helicase, two decades of studies on its structure function and regulation from its discovery opens a window for development of a non-hormonal oral male contraceptive. Front Endocrinol (Lausanne) 2019;10:576-586.
- Kavarthapu R, Anbazhagan R, Raju M, Morris CT, Pickel J, Dufau ML. Targeted knock-in mice with a human mutation in GRTH/DDX25 reveals the essential role of phosphorylated GRTH in spermatid development during spermatogenesis. Hum Mol Genet 2019;28:2561-2572.
- Raju M, Hassan SA, Kavarthapu R, Anbazhagan R, Dufau ML. Characterization of the phosphorylation site of GRTH/DDX25 and protein kinase A binding interface provides structural basis for the design of a non-hormonal male contraceptive. Sci Rep 2019;9:42857-42867.
- Kavarthapu R, Anbazhagan R, Sharma AK, Shiloach J, Dufau ML. Linking phospho-gonadotropin regulated testicular RNA helicase (GRTH/DDX25) to histone ubiquitination and acetylation essential for spermatid development during spermiogenesis. Front Cell Dev Biol 2020;8:310-321.
Collaborators
- Sergio A. Hassan, PhD, Center for Molecular Modeling, CIT, NIH, Bethesda, MD
Contact
For more information, email dufau@helix.nih.gov or visit http://irp.nih.gov/pi/maria-dufau.