Molecular Genetics of an Imprinted Gene Cluster on Mouse Distal Chromosome 7

- Karl Pfeifer, PhD, Head, Section on Epigenetics
- Claudia Gebert, PhD, Biologist
- Beenish Rahat, PhD, Postdoctoral Fellow
- Christopher Tracy, PhD, Postdoctoral Fellow
- Hyungchul Lee, PhD, Visiting Fellow
- Jacob Noeker, BS, Postbaccalaureate Fellow
Genomic imprinting is an unusual form of gene regulation by which an allele’s parental origin restricts allele expression. For example, almost all expression of the noncoding RNA tumor-suppressor gene H19 is from the maternal chromosome. In contrast, expression of the neighboring Insulin-like Growth Factor 2 gene (Igf2) is from the paternal chromosome. Imprinted genes are not randomly scattered throughout the chromosome but rather are localized in discrete clusters where monoallelic expression is regulated by a common cis-acting DNA regulatory element called the Imprinting Control Region (ICR). We study a cluster of imprinted genes on the distal end of mouse chromosome 7 (Figure 1). The syntenic region in humans (11p15.5) is highly conserved in gene organization and expression patterns. Imprinting of H19 and of Igf2 is regulated by the H19ICR, which is located just upstream of the H19 promoter. We showed that the molecular function of the H19ICR is to organize the region into alternative 3D structures. Upon maternal inheritance, the H19ICR is not methylated, binds to the transcriptional repressor CTCF, and organizes the locus into loop structures that bring the H19 promoter into contact with downstream enhancers but exclude the Igf2 promoter from these enhancer interactions. Upon paternal inheritance, the H19ICR is methylated and cannot bind to CTCF. Therefore, alternative loop structures form, which allow Igf2 promoters to interact with the shared enhancers while preventing H19 promoter/enhancer interactions. In humans, epigenetic mutations that disrupt H19ICR function result in loss of monoallelic expression. Mutations in the paternal H19ICR lead to loss of Igf2 expression and biallelic (2X) H19 expression and are associated with the Russell-Silver syndrome. Mutations in the maternal H19ICR lead to loss of H19 but biallelic (2x) Igf2 expression and are associated with the Beckwith-Wiedemann syndrome and several pediatric cancers. Our lab generated mouse models that phenocopy the human diseases, and our goal is to characterize the molecular defects associated with misimpression of Igf2/H19 and to understand how these molecular defects lead to disease and cancer. In particular, we strive to understand the role of development in disease progression. In addition, to the Igf2/H19 locus, we are completing analyses of mice carrying conditional mutations at the Casq2 locus to understand the role of development in cardiac disease progression.
Click image to view.
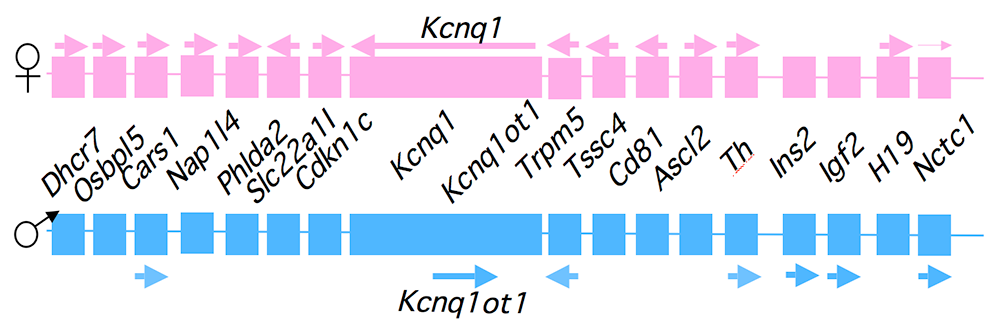
Figure 1. An imprinted domain on mouse distal chromosome 7
Maternal (pink) and paternal (blue) chromosomes are indicated. Horizontal arrows denote RNA transcription.
Alternative long-range interactions between distal regulatory elements establish allele-specific expression at the Igf2/H19 locus
Our studies on the mechanisms of genomic imprinting focus on the H19 and Igf2 genes. Paternally expressed Igf2 lies about 80 kb upstream of the maternal-specific H19 gene. Using cell-culture systems as well as transgene and knockout experiments in vivo, we identified the enhancer elements responsible for activation of the two genes. The elements are shared and are all located downstream of the H19 gene (Figure 2).
As mentioned above, imprinting at the Igf2/H19 locus depends on the 2.4 kb H19ICR, which lies between the two genes, just upstream of the H19 promoter (Figure 2). On the maternal chromosome, binding of the CTCF protein, a transcriptional regulator, to the H19ICR establishes a transcriptional insulator that organizes the chromosome into loops. The loops favor H19 expression but block interactions between the maternal Igf2 promoters and the downstream shared enhancers, thus preventing maternal Igf2 expression. Upon paternal inheritance, the cytosine residues within the ICR DNA sequences are methylated, which prevents binding of the CTCF protein, so that a transcriptional insulator is not established. Thus, paternal Igf2 promoters and the shared enhancers interact via DNA loops, and expression of paternal Igf2 is facilitated. Taken together, we find that the fundamental role of the ICR is to organize the chromosomes into alternative 3-D configurations that promote or prevent expression of the Igf2 and H19 genes.
Click image to view.
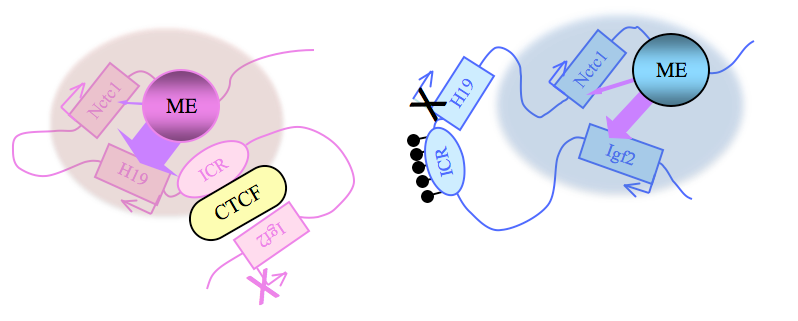
Figure 2. Distinct maternal and paternal chromosomal conformations at the distal 7 locus
Epigenetic modifications on the 2.4 kb ICR generate alternative 3D organizations across a large domain on paternal (blue) and maternal (pink) chromosomes and thereby regulate gene expression. ICR, imprinting control region; ME, muscle enhancer; filled lollipops, CpG methylation covering the paternal ICR.
The H19 ICR is not only necessary but is also sufficient for genomic imprinting. To demonstrate this, we used knock-in experiments to insert the 2.4 kb element at heterologous loci and demonstrated its ability to imprint these regions. Further, analyses of the loci confirmed and extended the transcriptional model described above. Upon maternal inheritance, even ectopic ICR elements remain unmethylated, bind to the CTCF protein, and form transcriptional insulators. Paternally inherited ectopic ICRs become methylated, cannot bind to CTCF, and therefore promote alternative loop domains distinct from those organized on maternal chromosomes. Most curious was the finding that DNA methylation of ectopic ICRs is not acquired until relatively late in development, after the embryo implants in the uterus. In contrast, at the endogenous locus, ICR methylation occurs during spermatogenesis. The findings thus imply that DNA methylation is not the primary imprinting mark that distinguishes maternally from paternally inherited ICRs.
The Nctc1 gene lies downstream of H19 and encodes a long noncoding RNA that is transcribed across the muscle enhancer element (ME in Figure 2), which is shared by Igf2 and H19. Nctc1 expression depends on this enhancer element. Concordantly, the shared enhancer interacts with the Nctc1 promoter, just as it interacts with the maternal H19 and paternal Igf2 promoters. We showed that all three co-regulated promoters (Igf2, H19, and Nctc1) also physically interact with each other in a manner that depends on their interactions with the shared enhancer. Thus, enhancer interactions with one promoter do not preclude interactions with another promoter. Moreover, we demonstrated that such promoter-promoter interactions are regulatory; they explain the developmentally regulated imprinting of Nctc1 transcription. Taken together, our results demonstrate the importance of long-range enhancer-promoter and promoter-promoter interactions in physically organizing the genome and establishing the gene expression patterns that are crucial for normal mammalian development.
Molecular mechanisms for tissue-specific promoter activation by distal enhancers
Normal mammalian development is absolutely dependent on establishing the appropriate patterns of expression of thousands of developmentally regulated genes. Most often, development-specific expression depends on promoter activation by distal enhancer elements. The Igf2/H19 locus is a highly useful model system for investigating mechanisms of enhancer activation. First, the biological significance of the model is clear, given that expression of these genes is so strictly regulated. Even twofold changes in RNA levels are associated with cancer and developmental disorders. Second, we already know much about the enhancers in this region and have established powerful genetic tools to investigate their function. Igf2 and H19 are co-expressed throughout embryonic development and depend on a series of tissue-specific enhancers that lie between 8 and more than 150 kb downstream of the H19 promoter (or between 88 and more than 130 kb downstream of the Igf2 promoters). The endodermal and muscle enhancers have been precisely defined, and we generated mouse strains carrying deletions that completely abrogate enhancer function. We also generated insulator insertion mutations that specifically block muscle enhancer activity. We used these strains to generate primary myoblast cell lines so that we can combine genetic, molecular, biochemical, and genomic analyses to understand the molecular bases for enhancer functions.
A long noncoding RNA is an essential element of the muscle enhancer
Transient transfection analyses define a 300–bp element that is both necessary and sufficient for maximal enhancer activity. However, stable transfection and mouse mutations indicate that this core element is not sufficient for enhancer function in a chromosomal context. Instead, the Nctc1 promoter element is also essential (Nctc1 encodes a spliced, polyadenylated long noncoding RNA); Nctc1 RNA itself is not required (at least in trans). Instead mutational analysis demonstrates that it is Nctc1 transcription through the core enhancer that is necessary for enhancer function. Curiously, the Nctc1 promoter has chromatin features typical of both a classic enhancer and a classic peptide-encoding promoter. Several recent genomic studies also suggested a role for noncoding RNAs in gene regulation and enhancer function. We will use our model system to characterize the role of Nctc1 transcription in establishing enhancer orientation, enhancer promoter specificity, and enhancer tissue specificity.
The muscle enhancer (ME) directs RNA polymerase (RNAP) II not only to its cognate promoters (i.e., to the H19 and Igf2 promoters) but also across the entire intergenic region.
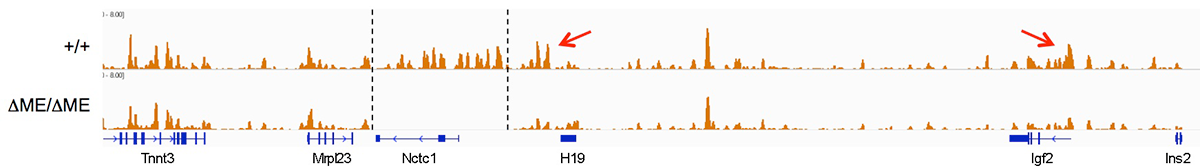
We used ChIP-on-chip to analyze RNAP localization on chromatin prepared from wild-type and from enhancer-deletion (DME) cell lines (Figure 3). As expected, RNAP binding to the H19 and Igf2 promoters is entirely enhancer-dependent. Curiously, we also noted enhancer-dependent RNAP localization across the entire locus, including the large intergenic domain between the two genes. Furthermore, RNAP binding is associated with RNA transcription. Thus, the enhancer regulates accessibility and RNAP binding not solely at specific localized sites but across the entire domain. The results support a facilitated tracking model for enhancer activity.
Click image to view.
Figure 3. The shared muscle enhancer (ME) directs RNAP binding and RNA transcription across the entire 150 kb locus.
RNAP binding at 'real' genes and across the intergenic regions is qualitatively different.
We used naturally occurring single-nucleotide polymorphisms (SNPs) to investigate allelic differences in binding of RNAP and activation of gene expression in wild-type cells and in cells carrying enhancer deletions or insulator insertion mutations. RNAP binding across the Igf2 and H19 genes is both enhancer-dependent and insulator-sensitive; that is, a functional insulator located between an enhancer and its regulated gene prevents RNAP binding and likewise prevents RNA transcription. Across the intergenic regions, RNAP binding and RNA transcription are similarly enhancer-dependent (see above). However, intergenic RNAP binding and transcription are not insulator-sensitive. The results indicate that insulators do not serve solely as a physical block for RNAP progression, but rather they specifically interfere with certain RNAP states or activities.
The muscle enhancer regulates RNAP binding and RNA transcription, but does not establish chromatin structures.
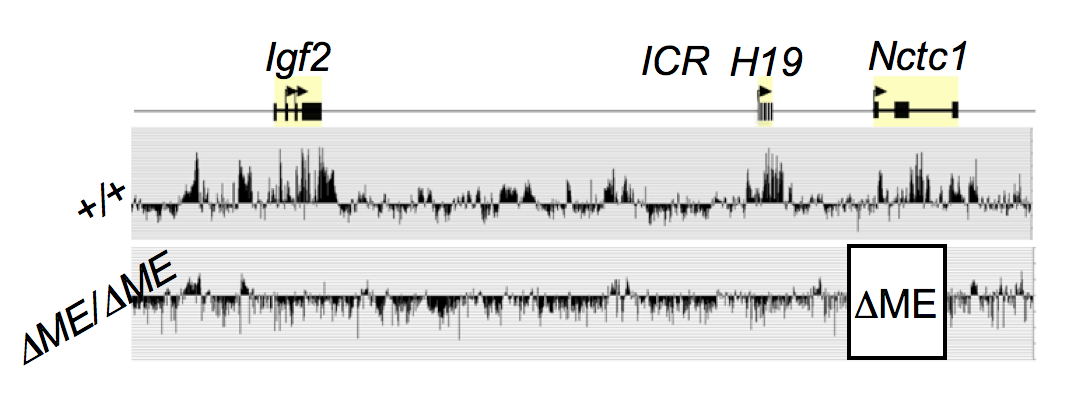
Both RNA transcription and RNAP binding across the Igf2/H19 domain are entirely dependent upon the muscle enhancer. For example, levels of H19 RNA are reduced more than 10,000-fold in muscle cells in which the enhancer has been deleted. To test the dependence of chromatin structure on enhancer activity, we performed ChiP-Seq on wild-type and on enhancer-deletion cell lines using antibodies to the histones H3K4me1, H3K43me3, and H3K36me3. Surprisingly, we saw no changes in the patterns of chromatin modification (Figure 4). Thus, a functional enhancer and active RNA transcription are not important for establishing chromatin structures at the Igf2/H19 domain.
Click image to view.
Figure 4. Chromatin patterns at the Igf2/H19 locus are independent of enhancer activity.
Chromatin was isolated from wild-type and enhancer-deletion muscle cells, using antibodies to H3K4me1, and analyzed by DNA sequencing.
Function of the H19 and Igf2 genes in muscle-cell growth and differentiation [Reference 1]
Misexpression of H19 and IGF2 is associated with several developmental diseases (including Beckwith-Wiedemann syndrome and Silver-Russell syndrome) and with several kinds of cancer, especially Wilms’ tumor and rhabdosarcoma. In humans, misexpression is most often caused by loss-of-imprinting mutations that result in biallelic expression of IGF2 and loss of expression of H19. We generated and characterized primary myoblast cell lines from mice carrying deletion of the H19 ICR that phenocopies the loss-of-imprinting expression phenotypes; that is, H19icr–deletion mice make extra Igf2 but no H19. Mice carrying this mutation do not develop rhabdosarcoma but show defects in their ability to respond to and to heal muscle injury. Moreover, primary myoblast lines derived from mutant mice are defective in their ability to differentiate in vitro (Figure 5).
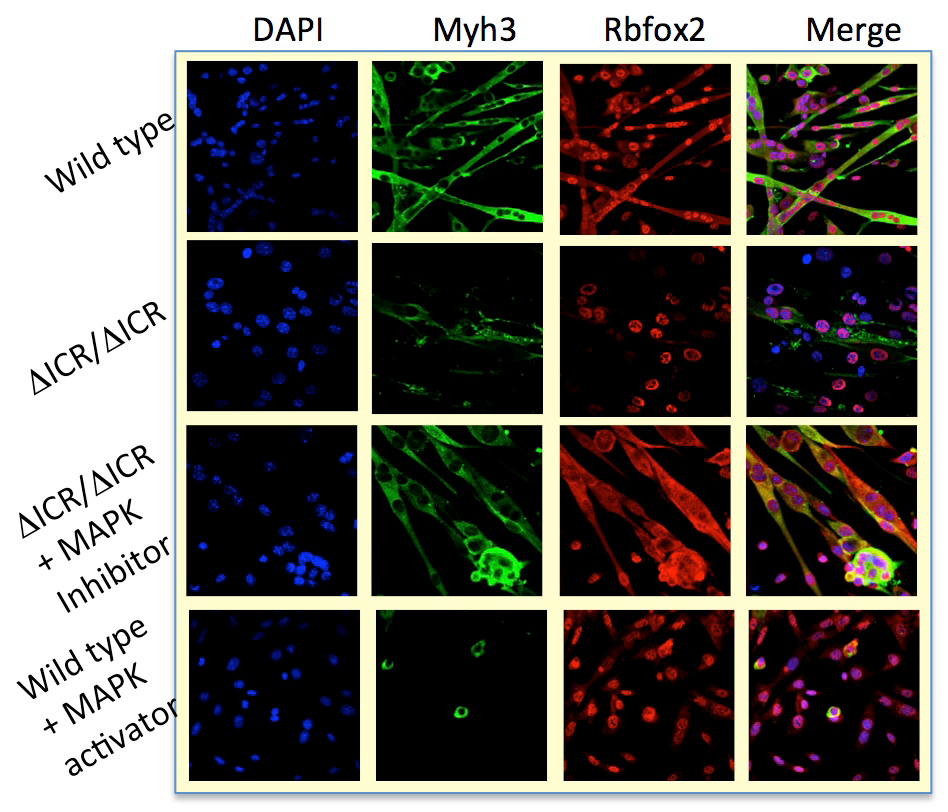
Click image to view.
Figure 5. Muscle cell–differentiation defects in Igf2/H19 loss-of-imprinting mice
Differentiation defects in loss-of-imprinting (ΔICR) myoblasts can be rescued by blocking MAP kinase 3 activity. Conversely, artificial activation of the MAPK activity in wild-type cells mimics the genetic defect.
To understand the molecular basis for the differentiation phenotype, we performed RNA sequencing and identified several hundred genes whose expression levels are altered by the ICR deletion. GO (gene ontology) pathway analysis demonstrates that these differentially expressed genes were highly enriched in the MAP kinase signaling pathway. Of special note, expression of the Mapk3 gene is elevated in mutant cell lines.
To determine the significance of the changes in Mapk3, we used drug inhibitors to block MAP kinase activity. In mutant cell lines, we can restore normal differentiation by blocking activation of the MAP kinase target MEK1. Similarly, treatments that activate MAP kinase in wild-type cells can mimic the ICR–deletion phenotype. The results suggest that H19/Igf2 act through MAP kinase to regulate differentiation of myoblast cells.
To distinguish the roles of Igf2 overexpression and H19 underexpression, we analyzed additional mouse strains that restore H19 via a bacterial artificial chromosome transgene or that restore normal levels of Igf2 expression via a second mutation in the paternal Igf2 gene. Analyses of cell lines from such mice demonstrate that extra Igf2 is the direct cause of failure to differentiate in loss-of-imprinting mutations, but that H19 is essential for normal fusion and for muscle hypertrophy in response to Wnt pathways (Figure 6). Molecular and genetic analyses indicate that H19 normally functions to bind to p53 and reduce its bioavailability. Thus, loss of H19 results in increased p53 function and therefore to enhanced activation of the mTOR/AKT signaling pathways.
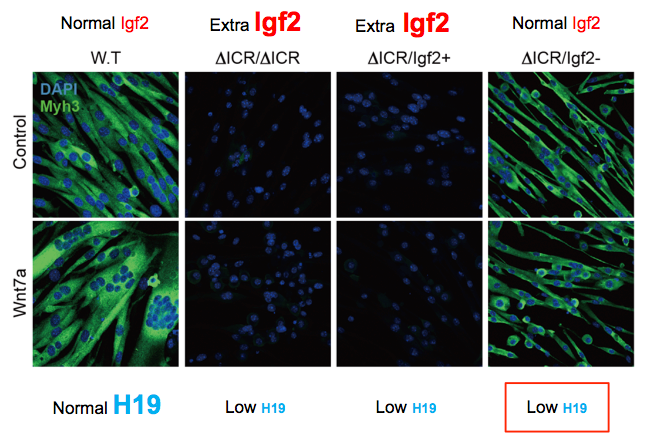
Click image to view.
Figure 6. The long noncoding H19 RNA is required for normal myotube fusion and hypertrophy.
Loss-of-imprinting defects at the Igf2/H19 locus result in extra expression of Igf2 and defects in myotube differentiation: Compare W.T (wild-type) with ΔICR/DICR and ΔICR/Igf2+ cells. Mutation of the paternal Igf2 gene can restore normal Igf2 expression levels and thus restore normal differentiation (see ΔICR/Igf2– cells). However, these cells still do not make the H19 long non-coding RNA, do not fuse efficiently, and do not respond to Wnt7a signaling.
Function of H19 and Igf2 genes in cardiac development
Beckwith-Wiedeman syndrome (BWS) is a developmental disorder characterized by generalized overgrowth of the fetus and a high risk for several neonatal cancers. Many BWS patients also display cardiac problems. BWS can be explained by one of two different genetic lesions: loss of function of the CDKN1C gene or maternal loss of imprinting at the H19/Igf2 locus. Maternal loss of imprinting has the effect of doubling Igf2 expression while concomitantly reducing H19 RNA levels. Curiously, children born via artificial reproductive technology (ART) show increased incidence of BWS, which can be explained by increasingly frequent loss of H19/Igf2 imprinting in these children. Moreover the children show high frequency of cardiac dysfunction. Altogether, these results suggest that abnormal expression of the H19/Igf2 locus can lead to cardiac problems.
We observed that our BWS mouse model also results in cardiac dysfunction, as measured by echocardiography and ECG analyses. Molecular and molecular-genetic analyses demonstrate that biallelic Igf2 and loss of H19 play independent and distinct roles in generating the BWS phenotype.
Biallelic expression of Igf2 results in increased levels of circulating IGF2 peptide, which superactivates insulin and insulin-like receptor kinases in cardiomyocytes, resulting in hyper-activation of AKT/mTOR signaling pathways, which in turn causes cardiomyocyte hypertrophy and hyperplasia. Such effects result in a cardiac hypertrophy that is non-pathologic and transient, that is, the hearts function normally and, as long as H19 levels are normal, the heart size normalizes after birth once Igf2 expression is repressed. Thus, there are no significant health effects associated with loss of imprinting of Igf2 only.
Loss of expression of H19 is pathologic. Hearts show progressive heart disease as manifested by hypertrophy, increased fibrosis, expression of cardiac failure markers, and reduced and abnormal heart function, as measured by echocardiography. In hearts, H19 expression is restricted to endothelial cells. In vivo analyses of whole hearts and in vitro analyses of isolated endothelial cells show that reduction in H19 results in increased endothelial-to-mesenchymal transition (EMT). EMT is a process that is an essential feature of normal cardiac development; for example, formation of cardiac valves requires EMT. However, elevated frequency of EMT is associated with heart disease. Our data support the idea that H19 regulates the cell fate of endothelial cells, and future experiments aim to identify the molecular mechanisms.
Role of calsequestrin2 in regulating cardiac function [Reference 3]
Mutations in the CASQ2 gene, which encodes cardiac calsequestrin (CASQ2), are associated with catecholaminergic polymorphic ventricular tachycardia (CPVT) and sudden death. The survival of individuals homozygous for loss-of-function mutations in CASQ2 was surprising, given the central role of Ca2+ ions in excitation-contraction (EC) coupling and the presumed critical roles of CASQ2 in regulating Ca2+ release from the sarcoplasmic reticulum (SR) into the cytoplasm. To address this paradox, we generated a mouse model for loss of Casq2 gene activity. Comprehensive analysis of cardiac function and structure yielded several important insights into CASQ2 function. First, CASQ2 is not essential to provide sufficient Ca2+ storage in the SR of the cardiomyocyte. Rather, a compensatory increase in SR volume and surface area in mutant mice appears to maintain normal Ca2+ storage capacity. Second, CASQ2 is not required for the rapid, triggered release of Ca2+ from the SR during cardiomyocyte contraction. Rather, the RyR receptor, an intracellular calcium-ion channel, opens appropriately, resulting in normal, rapid flow of Ca2+ into the cytoplasm, thus allowing normal contraction of the cardiomyocyte. Third, CASQ2 is required for normal function of the RyR during cardiomyocyte relaxation. In the absence of CASQ2, significant Ca2+ leaks occur through the RyR and lead to premature contractions and cardiac arrhythmias (Figure 7). Fourth, CASQ2 function is required to maintain normal levels of the SR proteins junctin and triadin. We do not yet understand what role, if any, the compensatory changes in these two SR proteins play in modulating the loss of Casq2 phenotype.
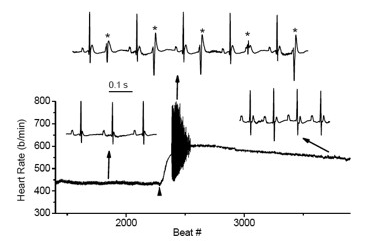
Click image to view.
Figure 7. Cardiac arrhythmias in calsequestrin-2–deficient mice phenocopy the human disease.
Premature ventricular complexes (*) are induced by stress in Casq2–deficient but not in wild-type mice.
To address these issues and to model cardiac disorders associated with late-onset (not congenital) loss of CASQ2 activity, we established and are analyzing two new mouse models in which changes in Casq2 gene structure are induced by tissue-specific transgenes activated by tamoxifen treatment. In the first model, an invested/null allele is restored to normal function by the addition of the drug. In the past year, we demonstrated the effectiveness of this model and noted that full CASQ2 protein levels are restored within one week of treatment. In the second model, a functional gene is ablated by the addition of the drug. The Casq2 gene and mRNAs are deleted from cardiac cells within four days of hormone treatment. Phenotypic analyses shows that restoration of CASQ2 in adult animals is sufficient to fully restore cardiac function. Moreover, restoration solely in pacemaking cells is also enough to rescue function, suggesting an important role for reduced heart rate in the CPVT phenotype as well as a new target for therapeutic interventions.
Publications
- Park K-S, Mitra A, Rahat B, Kim B, Pfeifer K. Loss of imprinting mutations define both distinct and overlapping roles for misexpression of IGF2 and of H19 lncRNA. Nucleic Acids Res 2017;45:12766-12779.
- Gebert C, Correia L, Li Z, Petrie HT, Love PE, Pfeifer K. Chromosome choice for initiation of V-(D)-J recombination is not governed by genomic imprinting. Immunol Cell Biol 2017;95:473-477.
- Flores DJ, Duong T, Brandenberger LO, Mitra A, Shirali A, Johnson JC, Springer D, Noguchi A, Yu ZX, Ebert SN, Ludwig A, Knollmann BC, Levin MD, Pfeifer, K. Conditional ablation and conditional rescue models for Casq2 elucidate the role of development and of cell-type specific expression of Casq2 in the CPVT2 phenotype. Hum Mol Genet 2018;27:1533-1544.
- Mitra A, Park K-S, Pfeifer K. Circular RNAs and competing endogenous (ceRNA) networks. Transl Cancer Res 2018;7:S624-628.
Collaborators
- Bjorn Knollmann, MD, PhD, Vanderbilt University Medical Center, Nashville, TN
- Mark D. Levin, MD, Cardiovascular & Pulmonary Branch, NHLBI, Bethesda, MD
- Danielle A. Springer, VMD, Dipl ACLAM, Animal Program, NHLBI, Bethesda, MD
Contact
For more information, email kpfeifer@helix.nih.gov or visit http://pfeiferlab.nichd.nih.gov.