Neuronal Circuits Controlling Behavior: Genetic Analysis in Zebrafish

- Harold Burgess, PhD, Head, Section on Behavioral Neurogenetics
- Tripti Gupta, PhD, Staff Scientist
- Ashwin A. Bhandiwad, PhD, Postdoctoral Fellow
- Eric J. Horstick, PhD, Postdoctoral Fellow
- Abhignya Subedi, PhD, Postdoctoral Fellow
- Kathryn M. Tabor, PhD, Postdoctoral Fellow
- Jennifer L. Strykowski, MSc, Zebrafish Technician
- Gregory Marquart, BSc, Graduate Student
- Yared Bayleyen, BSc, Postbaccalaureate Fellow
- Trevor S. Smith, BSc, Postbaccalaureate Fellow
Our goal is to understand how, under diverse environmental contexts, the nervous system selects appropriate behavioral actions in a way that best satisfies internal motivational objectives. We use the zebrafish as a model because, in the larval stage, its brain exhibits the basic architecture of the vertebrate brain but is much less complex than the mammalian brain. Despite the relative simplicity of their nervous system, larvae have a sophisticated repertoire of sensory-guided and internally driven behaviors. Furthermore, the optical clarity of the embryo facilitates visualization of individual neurons and their manipulation with genetic techniques. Behavior in larvae is innate and therefore exhibits minimal variability between fish. Subtle alterations in behavior can therefore be robustly scored, making it possible to quickly assess the contribution of identified neurons to a variety of motor behaviors.
We focus on two aspects of behavioral regulation: the mechanisms by which sensory context regulates behavioral decisions (Reference 1) and pathways that sustain changes in behavioral state (References 2,3). In addition, we are developing a suite of genetic tools and behavioral assays to probe the nexus between neuronal function and behavior at single-cell resolution (References 4,5). The neuronal connections that allow the brain to integrate sensory and internal-state information are established through genetic interactions during development. We aim to identify genes and neurons that are required for the functional development of such connections. In vertebrates, neuronal circuits situated in the brainstem form the core of the locomotor control network and are responsible for balance, posture, motor control, and arousal. Accordingly, many neurological disorders stem from abnormal formation or function of brainstem circuits. Insights into the function of brainstem circuits in health and disease have come from genetic manipulation of neurons in zebrafish larvae in combination with computational analysis of larval behavior.
Molecular identification of neurons that mediate prepulse inhibition
Startle responses are rapid reflexes that are triggered by sudden sensory stimuli and that help animals defend against or escape from potentially threatening stimuli. In both fish and mammals, startle responses are initiated by giant reticulospinal neurons in the medulla, which receive short-latency sensory input from diverse sensory modalities. Although highly stereotyped, startle responses are nevertheless modulated by sensory context and behavioral state and are therefore an excellent system in which to understand how such information is integrated for behavioral choice. In mammals, including humans, the startle response to a strong auditory stimulus can be inhibited by pre-exposure to a weak acoustic ‘prepulse.’ This form of startle modulation, termed prepulse inhibition (PPI), is diminished in several neurological conditions. Vibrational stimuli trigger rapid-escape swims in zebrafish, which are mediated by giant reticulospinal neurons, in a manner similar to the central neurons controlling startle responses in mammals. Escape swims are suppressed by pre-exposure to a prepulse, allowing us to apply the powerful suite of genetic tools available in zebrafish to identify neurons that mediate prepulse inhibition.
To identify a transgenic zebrafish line that genetically labels neurons required for PPI, we screened a library of neuron-specific Gal4–enhancer trap lines marking distinct populations of neurons in the brain, by ablating the neurons in each enhancer trap line before testing for PPI. The transgenic line y252 labels a discrete population of neurons in the hindbrain whose ablation or opto-genetic inhibition eliminates PPI. The neurons genetically labeled in y252 are specified by the transcription factor Gsx1. We found that Gsx1–knockout mice showed a strong reduction in PPI, suggesting that a conserved circuit involving Gsx1–specified neurons mediates PPI across vertebrate species (Reference 1). To identify the precise subset of Gsx1 neurons that mediate PPI, we then used volumetric calcium imaging to simultaneously visualize the activity in thousands of Gsx1 neurons during a PPI paradigm. At the same time, we recorded tail movement responses to startle stimuli, with and without prepulse exposure. The method enabled us to locate a specific region that contains Gsx1 neurons whose activity correlates strongly with behavioral PPI. To demonstrate that such neurons are causally related to PPI, we established an intersectional genetic method to ablate subgroups of Gsx1 neurons by developing a library of Cre lines (Figure 1) and a UAS vector that requires both Gal4 and Cre expression in the same cell for reporter expression (UAS:KillSwitch). Ablation of Gsx1 neurons in the PPI–active zone eliminated PPI. We next visualized the morphology and projections of these neurons in order to reveal how they connect to the core startle circuit. For this, we developed a new method that exploits the low efficiency of B3 recombinase in zebrafish to achieve stochastic labeling of isolated neurons in cells that co-express Gal4 and Cre recombinase. Gsx1 neurons that mediate PPI project to the lateral dendrite of the Mauthner cell, the command neuron for startle responses in fish, enabling them to gate auditory information. Given that PPI is abnormal in neuro-psychiatric disorders with developmental origins, including schizophrenia and autism, our work will help identify and probe fundamental defects in circuitry that are abnormal in these conditions.
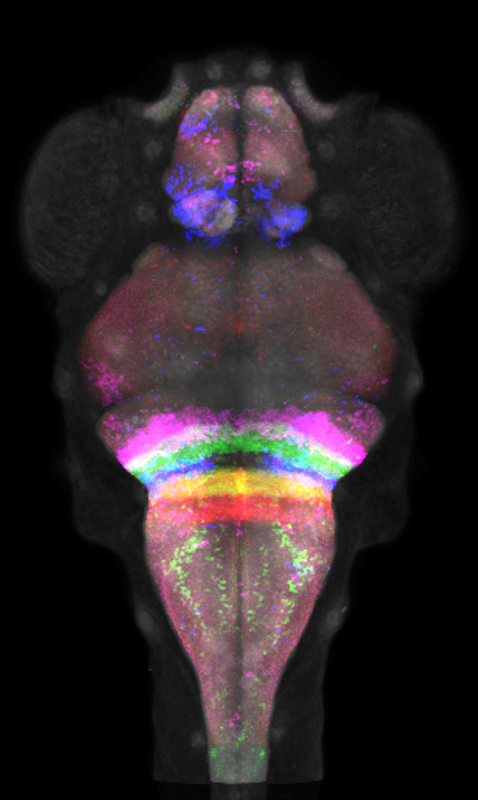
Click image to enlarge.
Figure 1. Intersectional interrogation of hindbrain circuitry using rhombomere-specific Cre lines
We developed a library of Cre lines that can be used in intersectional genetic experiments. Visualized here are confocal images of five Cre lines after co-registration to our atlas of transgene expression. Together, the lines partition the hindbrain into its rhombomeres 2–6, enabling neurons in each rhombomere to be functionally addressed.
Neural mechanisms for behavioral state control
Over the course of the day, motivational goals change in response to both internal and external cues. At any given moment, an individual's current behavioral state strongly influences decisions on how to interact with the environment. A major question in neuroscience is to identify the neural systems that maintain short-term behavioral states and to determine how they interact with central mechanisms for behavioral choice (Reference 2). In zebrafish, loss of illumination triggers a short-term behavioral state in which larvae show heightened locomotor activity. We previously demonstrated that light-sensitive neurons in the hypothalamus trigger this state of hyperactivity. However, the experiments were performed on larvae confined to small shallow chambers, making it difficult to characterize the underlying behavioral state. By recording larval movement in large-volume arenas, we found that the response to loss of illumination is partly light-search behavior. For the first two minutes in the dark, larval swim patterns are similar to a behavior characterized as area-restricted search across species, with high fractal dimension and repeated turning behavior (Figure 2). Larvae quickly locate and swim toward a light-spot if activated during this period. If no light-spot is found, the locomotor profile transitions to a remote-search behavior, in which larvae efficiently locate illuminated regions that are not visible in the original environment. Using a G0 cas9/crispr screen, we found that the transition between these two behavioral states requires the activity of otpa neurons, including opn4a–expressing deep brain photoreceptors (Reference 3).
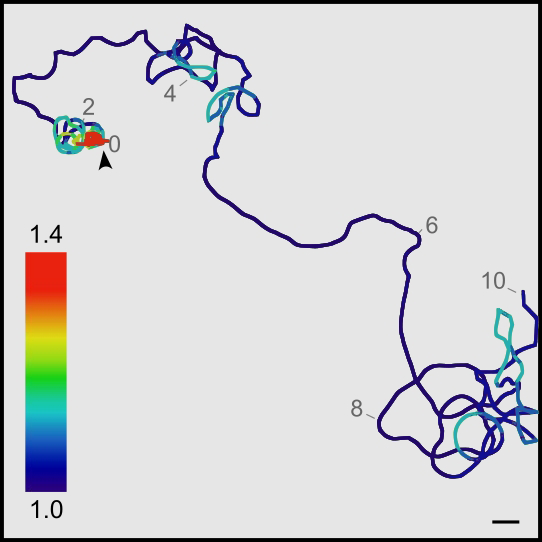
Click image to enlarge.
Figure 2. Local search behavior in larval zebrafish: swim trajectory of a single larva over a 10-minute period
The color scale indicates the fractal dimension within 30-second windows. Note that for the first two minutes, larvae swim in an area-restricted pattern, which then transitions to an outward-swim profile.
Tools for decoding neuronal circuits
A unique feature of brain imaging in zebrafish is the ability to visualize the total architecture of the brain while simultaneously recording the position and morphology of every constituent labeled neuron. Brain registration techniques enable data from several individuals to be quantitatively compared, so that experiments can systematically address the functional contributions of neurons across the entire brain. However, a challenge for deformable brain registration in larval zebrafish is to achieve highly precise global registration, without severely distorting the shape of individual cells. We calibrated parameters for the symmetric diffeomorphic normalization algorithm in ANTs (Advanced Normalization Tools) and found that it was possible to align larval zebrafish brains to a precision of around 1 cell diameter without sacrificing cell morphology. Using this method, we co-aligned more than 250 transgene expression patterns, derived from more than 1000 brain scans, to a single reference brain. Our Zebrafish Brain Browser (ZBB) software allows the expression in any of these lines to be compared at cellular-level resolution. Users can select a small region of the larval zebrafish brain and identity transgenic lines that express Gal4 or Cre in the corresponding region. Together with new tools that allow selective visualization or ablation of neurons that express both Gal4 and Cre, the ZBB atlas enables us to target highly specific neurons within the larval brain for visualizing activity or altering function, helping us and others decode their role within neuronal circuits (Reference 4). Moreover, multi-channel registration with ANTs allowed us to integrate our database of transgene expression patterns with Z-Brain, an independent zebrafish brain atlas that contains additional gene-expression patterns and neuro-anatomical annotations (Reference 5). To improve 3D visualization, we also developed tools for virtual reality projection of the zebrafish brain (Figure 3).

Click image to enlarge.
Figure 3. Virtual-reality zebrafish brain atlas
We developed a virtual-reality brain atlas for larval zebrafish. Using an inexpensive Cardboard viewer, different brain regions can be selected, enabling the neuro-anatomical structure of the larval brain to be visualized in three dimensions.
Additional Funding
- US Israel Binational Fund 2013433 (2014): Ongoing, with Yoav Gothilf, Tel Aviv University
Publications
- Bergeron SA, Carrier N, Li GH, Ahn S, Burgess HA. Gsx1 expression defines neurons required for prepulse inhibition. Mol Psychiatry 2015 20:974-85.
- Horstick EJ, Mueller T, Burgess HA. Motivated state control in larval zebrafish: behavioral paradigms and anatomical substrates. J Neurogenet 2016 30:122-132.
- Horstick EJ, Bayleyen Y, Sinclair JL, Burgess HA. Search strategy is regulated by somatostatin signaling and deep brain photoreceptors in zebrafish. BMC Biology 2017 15:4.
- Horstick EJ, Tabor KM, Jordan DC, Burgess HA. Genetic silencing and selective activation using nitroreductase and TTX insensitive channels. Methods Mol Biol: Zebrafish 2016 1451:355-361.
- Marquart GD, Tabor KM, Horstick EJ, Brown M, Burgess HA. High precision registration between zebrafish brain atlases using symmetric diffeomorphic normalization. GigaScience 2017 6:1-15.
Collaborators
- Kevin Briggman, PhD, Circuit Dynamics and Connectivity Unit, NINDS, Bethesda, MD
- Igor Dawid, PhD, Section on Developmental Biology, NICHD, Bethesda, MD
- Yoav Gothilf, PhD, Tel-Aviv University, Tel-Aviv, Israel
- Ralph Nelson, PhD, Basic Neurosciences Program, NINDS, Rockville, MD
- Sinisa Pajevic, PhD, Mathematical and Statistical Computing Lab, CIT, NIH, Bethesda, MD
Contact
For more information, email haroldburgess@mail.nih.gov or visit http://ubn.nichd.nih.gov.